

Short CDRL1 in intermediate VRC01-like mAbs is not sufficient to overcome key glycan barriers on HIV-1 Env
- PMID: 39240111
- PMCID: PMC11495006
- DOI: 10.1128/jvi.00744-24
Short CDRL1 in intermediate VRC01-like mAbs is not sufficient to overcome key glycan barriers on HIV-1 Env
Abstract
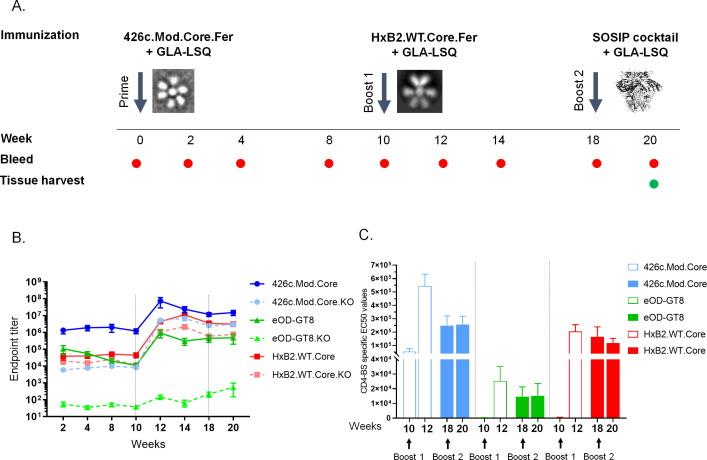
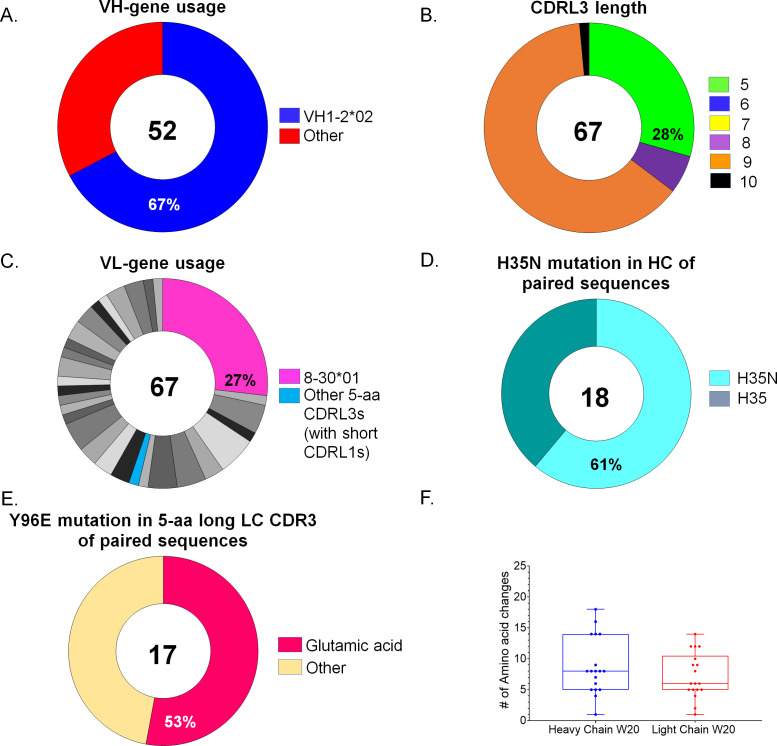
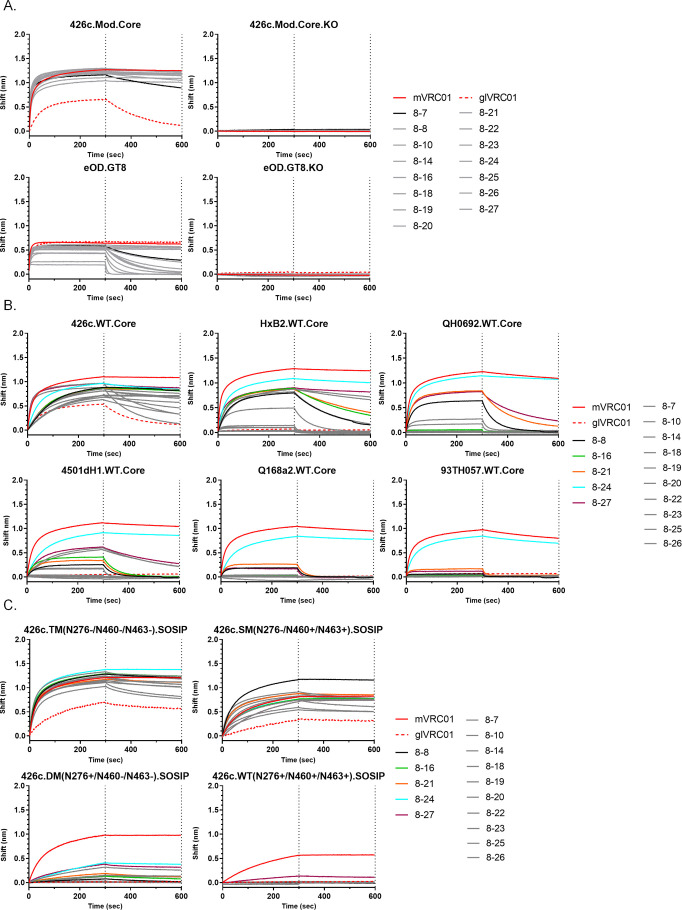
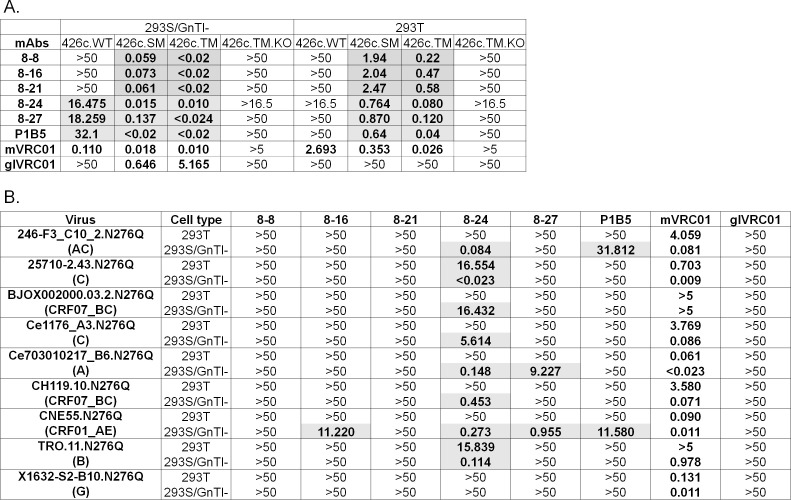
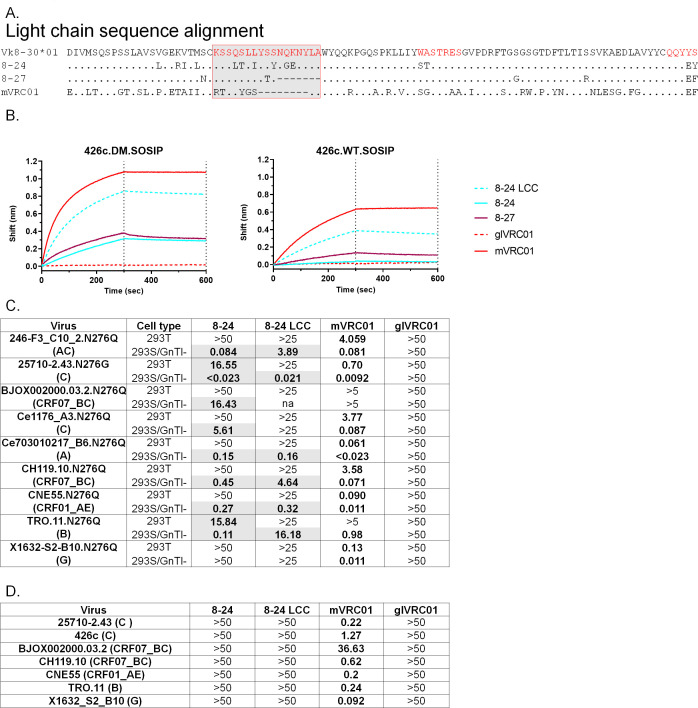
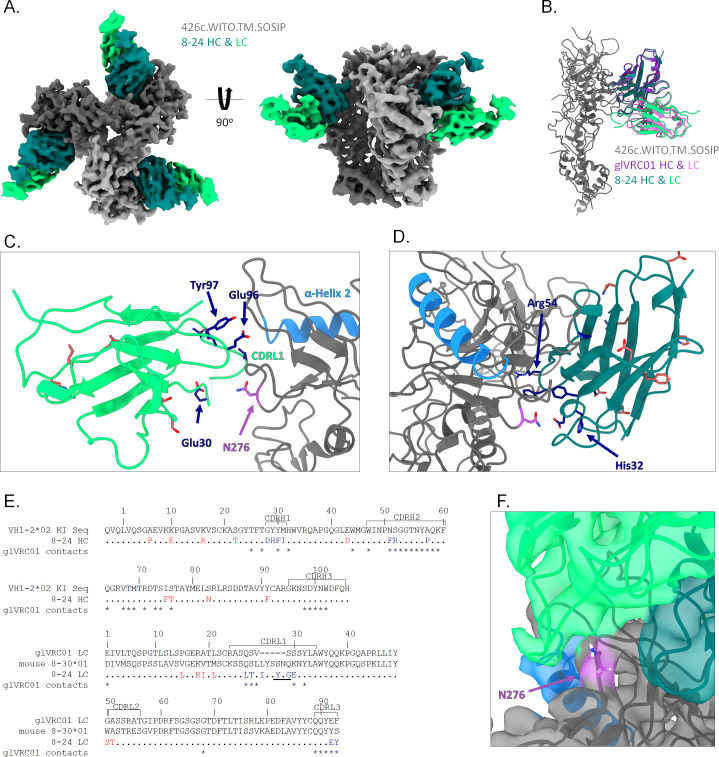
VRC01-class broadly neutralizing antibodies (bnAbs) have been isolated from people with HIV-1, but they have not yet been elicited by vaccination. They are extensively somatically mutated and sometimes accumulate CDRL1 deletions. Such indels may allow VRC01-class antibodies to accommodate the glycans expressed on a conserved N276 N-linked glycosylation site in loop D of the gp120 subunit. These glycans constitute a major obstacle in the development of VRC01-class antibodies, as unmutated antibody forms are unable to accommodate them. Although immunizations of knock-in mice expressing human VRC01-class B-cell receptors (BCRs) with specifically designed Env-derived immunogens lead to the accumulation of somatic mutations in VRC01-class BCRs, CDRL1 deletions are rarely observed, and the elicited antibodies display narrow neutralizing activities. The lack of broad neutralizing potential could be due to the absence of deletions, the lack of appropriate somatic mutations, or both. To address this point, we modified our previously determined prime-boost immunization with a germline-targeting immunogen nanoparticle (426c.Mod.Core), followed by a heterologous core nanoparticle (HxB2.WT.Core), by adding a final boost with a cocktail of various stabilized soluble Env trimers. We isolated VRC01-like antibodies with extensive somatic mutations and, in one case, a seven-amino acid CDRL1 deletion. We generated chimeric antibodies that combine the vaccine-elicited somatic mutations with CDRL1 deletions present in human mature VRC01 bnAbs. We observed that CDRL1 indels did not improve the neutralizing antibody activities. Our study indicates that CDRL1 length by itself is not sufficient for the broadly neutralizing phenotype of this class of antibodies.
Importance: HIV-1 broadly neutralizing antibodies will be a key component of an effective HIV-1 vaccine, as they prevent viral acquisition. Over the past decade, numerous broadly neutralizing antibodies (bnAbs) have been isolated from people with HIV. Despite an in-depth knowledge of their structures, epitopes, ontogenies, and, in a few rare cases, their maturation pathways during infection, bnAbs have, so far, not been elicited by vaccination. This necessitates the identification of key obstacles that prevent their elicitation by immunization and overcoming them. Here we examined whether CDRL1 shortening is a prerequisite for the broadly neutralizing potential of VRC01-class bnAbs, which bind within the CD4 receptor binding site of Env. Our findings indicate that CDRL1 shortening by itself is important but not sufficient for the acquisition of neutralization breadth, and suggest that particular combinations of amino acid mutations, not elicited so far by vaccination, are most likely required for the development of such a feature.
Keywords: BCR sequencing; CDRL1; HIV-1; VRC01-class antibodies; neutralization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
HIV-1 envelope glycan modifications that permit neutralization by germline-reverted VRC01-class broadly neutralizing antibodies.PLoS Pathog. 2018 Nov 5;14(11):e1007431. doi: 10.1371/journal.ppat.1007431. eCollection 2018 Nov. PLoS Pathog. 2018. PMID: 30395637 Free PMC article.
-
Inference of the HIV-1 VRC01 Antibody Lineage Unmutated Common Ancestor Reveals Alternative Pathways to Overcome a Key Glycan Barrier.Immunity. 2018 Dec 18;49(6):1162-1174.e8. doi: 10.1016/j.immuni.2018.10.015. Epub 2018 Dec 11. Immunity. 2018. PMID: 30552024 Free PMC article.
-
Overcoming Steric Restrictions of VRC01 HIV-1 Neutralizing Antibodies through Immunization.Cell Rep. 2019 Dec 3;29(10):3060-3072.e7. doi: 10.1016/j.celrep.2019.10.071. Cell Rep. 2019. PMID: 31801073 Free PMC article.
-
HIV Broadly Neutralizing Antibodies: VRC01 and Beyond.Adv Exp Med Biol. 2018;1075:53-72. doi: 10.1007/978-981-13-0484-2_3. Adv Exp Med Biol. 2018. PMID: 30030789 Review.
-
HIV-1 immunogens and strategies to drive antibody responses towards neutralization breadth.Retrovirology. 2018 Nov 26;15(1):74. doi: 10.1186/s12977-018-0457-7. Retrovirology. 2018. PMID: 30477581 Free PMC article. Review.
Cited by
-
Priming VRC01-precursor B cells with non-envelope immunogens disfavors boosting with HIV-1 envelope.NPJ Vaccines. 2025 Aug 5;10(1):185. doi: 10.1038/s41541-025-01235-5. NPJ Vaccines. 2025. PMID: 40764319 Free PMC article.
-
Increased immunogen valency improves the maturation of vaccine-elicited HIV-1 VRC01-class antibodies.bioRxiv [Preprint]. 2025 Mar 14:2025.03.13.642975. doi: 10.1101/2025.03.13.642975. bioRxiv. 2025. Update in: PLoS Pathog. 2025 May 29;21(5):e1013039. doi: 10.1371/journal.ppat.1013039. PMID: 40161829 Free PMC article. Updated. Preprint.
-
Increased immunogen valency improves the maturation of vaccine-elicited HIV-1 VRC01-like antibodies.PLoS Pathog. 2025 May 29;21(5):e1013039. doi: 10.1371/journal.ppat.1013039. eCollection 2025 May. PLoS Pathog. 2025. PMID: 40440315 Free PMC article.
References
-
- Barnes CO, Schoofs T, Gnanapragasam PNP, Golijanin J, Huey-Tubman KE, Gruell H, Schommers P, Suh-Toma N, Lee YE, Cetrulo Lorenzi JC, Piechocka-Trocha A, Scheid JF, West AP, Walker BD, Seaman MS, Klein F, Nussenzweig MC, Bjorkman PJ. 2022. A naturally arising broad and potent CD4-binding site antibody with low somatic mutation. Sci Adv 8:eabp8155. doi:10.1126/sciadv.abp8155 - DOI - PMC - PubMed
-
- Sajadi MM, Dashti A, Rikhtegaran Tehrani Z, Tolbert WD, Seaman MS, Ouyang X, Gohain N, Pazgier M, Kim D, Cavet G, Yared J, Redfield RR, Lewis GK, DeVico AL. 2018. Identification of near-pan-neutralizing antibodies against HIV-1 by deconvolution of plasma humoral responses. Cell 173:1783–1795. doi:10.1016/j.cell.2018.03.061 - DOI - PMC - PubMed
-
- Scheid JF, Mouquet H, Ueberheide B, Diskin R, Klein F, Oliveira TYK, Pietzsch J, Fenyo D, Abadir A, Velinzon K, Hurley A, Myung S, Boulad F, Poignard P, Burton DR, Pereyra F, Ho DD, Walker BD, Seaman MS, Bjorkman PJ, Chait BT, Nussenzweig MC. 2011. Sequence and structural convergence of broad and potent HIV antibodies that mimic CD4 binding. Science 333:1633–1637. doi:10.1126/science.1207227 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
