

MYC Drives mRNA Pseudouridylation to Mitigate Proliferation-Induced Cellular Stress during Cancer Development
- PMID: 39240686
- PMCID: PMC11609627
- DOI: 10.1158/0008-5472.CAN-24-1102
MYC Drives mRNA Pseudouridylation to Mitigate Proliferation-Induced Cellular Stress during Cancer Development
Abstract
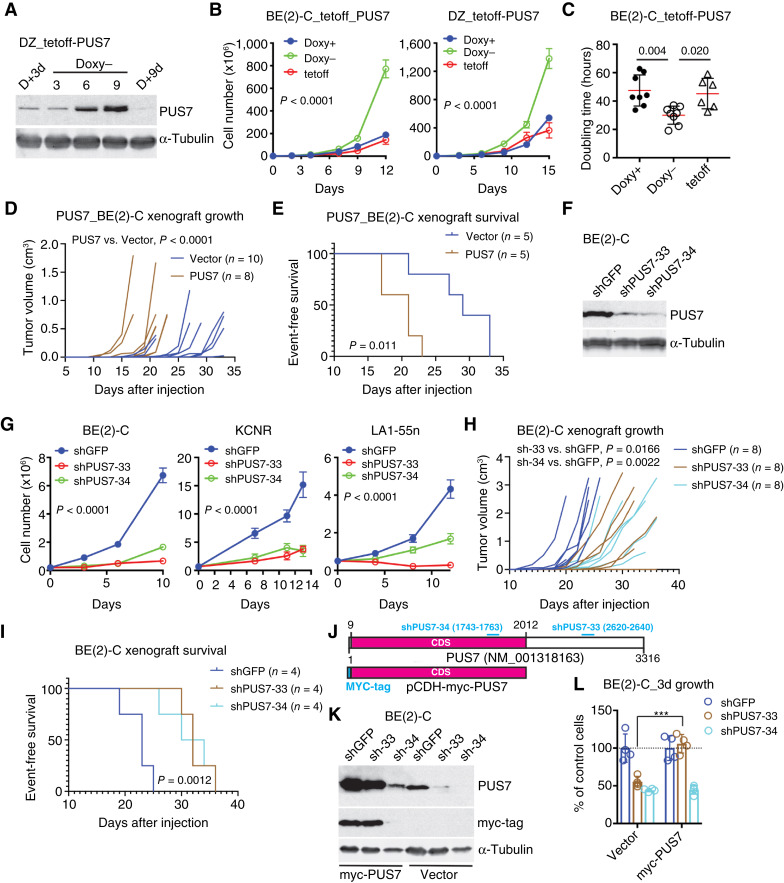
Pseudouridylation is a common RNA modification that is catalyzed by the family of pseudouridine synthases (PUS). Pseudouridylation can increase RNA stability and rigidity, thereby impacting RNA splicing, processing, and translation. Given that RNA metabolism is frequently altered in cancer, pseudouridylation may be a functionally important process in tumor biology. Here, we show that the MYC family of oncoproteins transcriptionally upregulates PUS7 expression during cancer development. PUS7 is essential for the growth and survival of MYC-driven cancer cells and xenografts by promoting adaptive stress responses and amino acid biosynthesis and import. ATF4, a master regulator of stress responses and cellular metabolism, was identified as a key downstream mediator of PUS7 functional activity. Induction of ATF4 by MYC oncoproteins and cellular stress required PUS7, and ATF4 overexpression overcame the growth inhibition caused by PUS7 deficiency. Mechanistically, PUS7 induced pseudouridylation of MCTS1 mRNA, which enhanced its translation. MCTS1, a noncanonical translation initiation factor, drove stress-induced ATF4 protein expression. A PUS7 consensus pseudouridylation site in the 3' untranslated region of ATF4 mRNA was crucial for the induction of ATF4 by cellular stress. These findings unveil an MYC-activated mRNA pseudouridylation program that mitigates cellular stress induced by MYC stimulation of proliferation and biomass production, suggesting that targeting PUS7 could be a therapeutic strategy selectively against MYC-driven cancers. Significance: Oncogene activation of mRNA pseudouridylation is a mechanism that facilitates metabolic reprogramming and adaptive responses to overcome cellular stress during cancer development.
©2024 The Authors; Published by the American Association for Cancer Research.
Conflict of interest statement
S. Sudarshan reports grants from NIH and the Department of Veterans Affairs during the conduct of the study. H.-F. Ding reports grants from NIH during the conduct of the study. No disclosures were reported by the other authors.
Figures







References
-
- Delaunay S, Helm M, Frye M. RNA modifications in physiology and disease: towards clinical applications. Nat Rev Genet 2024;25:104–22. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
