

Convergent evolution of monocyte differentiation in adult skin instructs Langerhans cell identity
- PMID: 39241057
- PMCID: PMC7616733
- DOI: 10.1126/sciimmunol.adp0344
Convergent evolution of monocyte differentiation in adult skin instructs Langerhans cell identity
Abstract
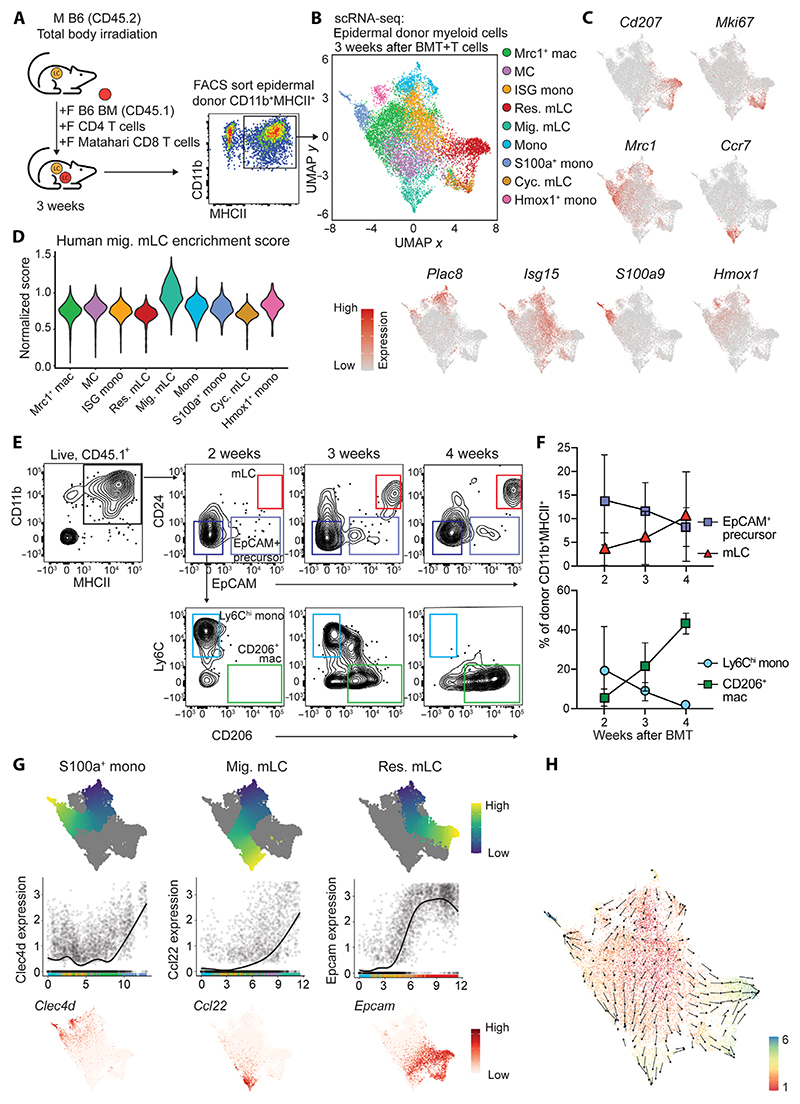
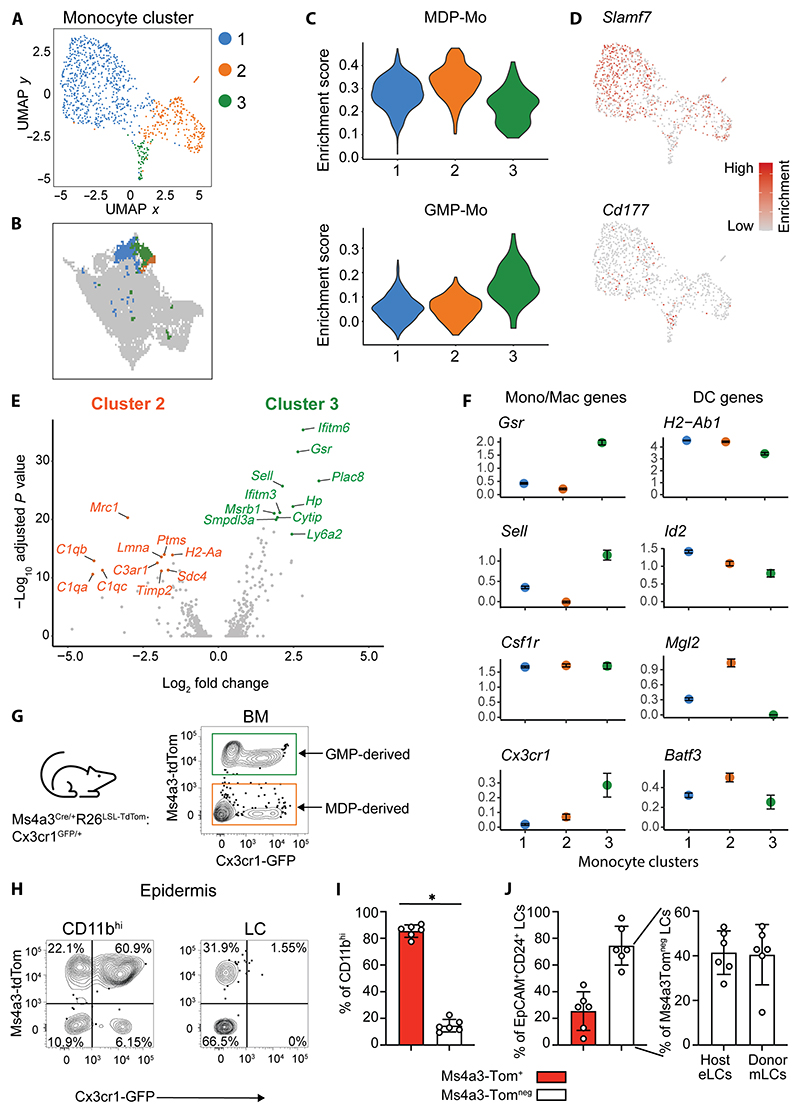
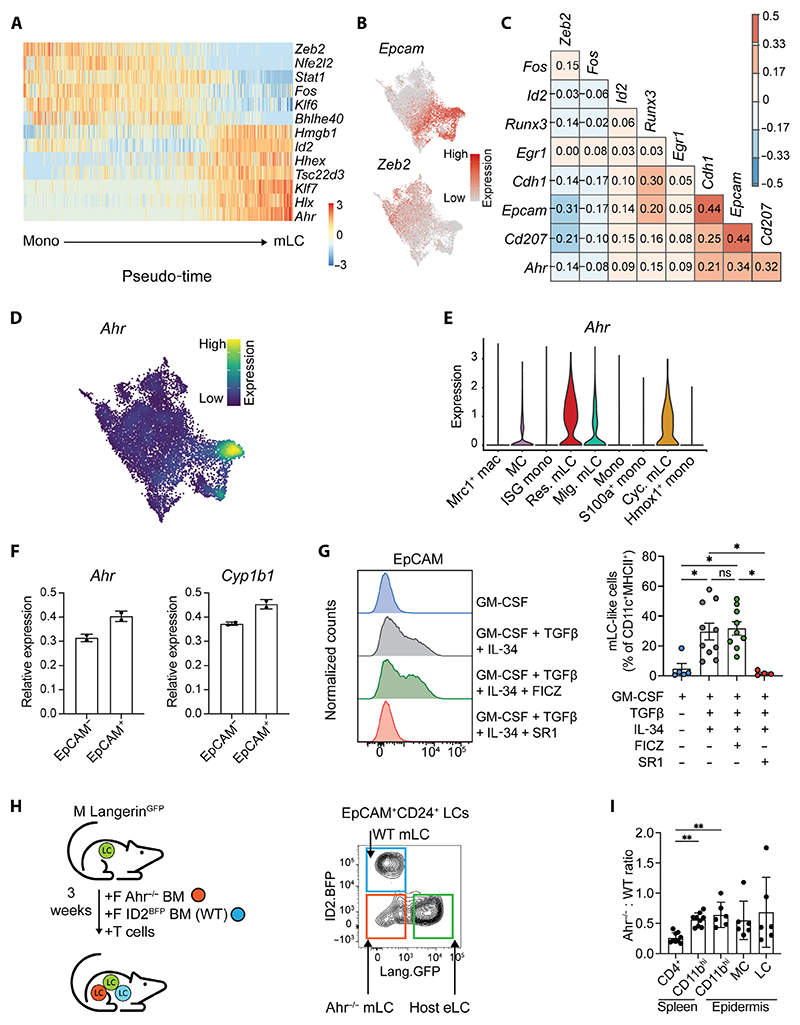
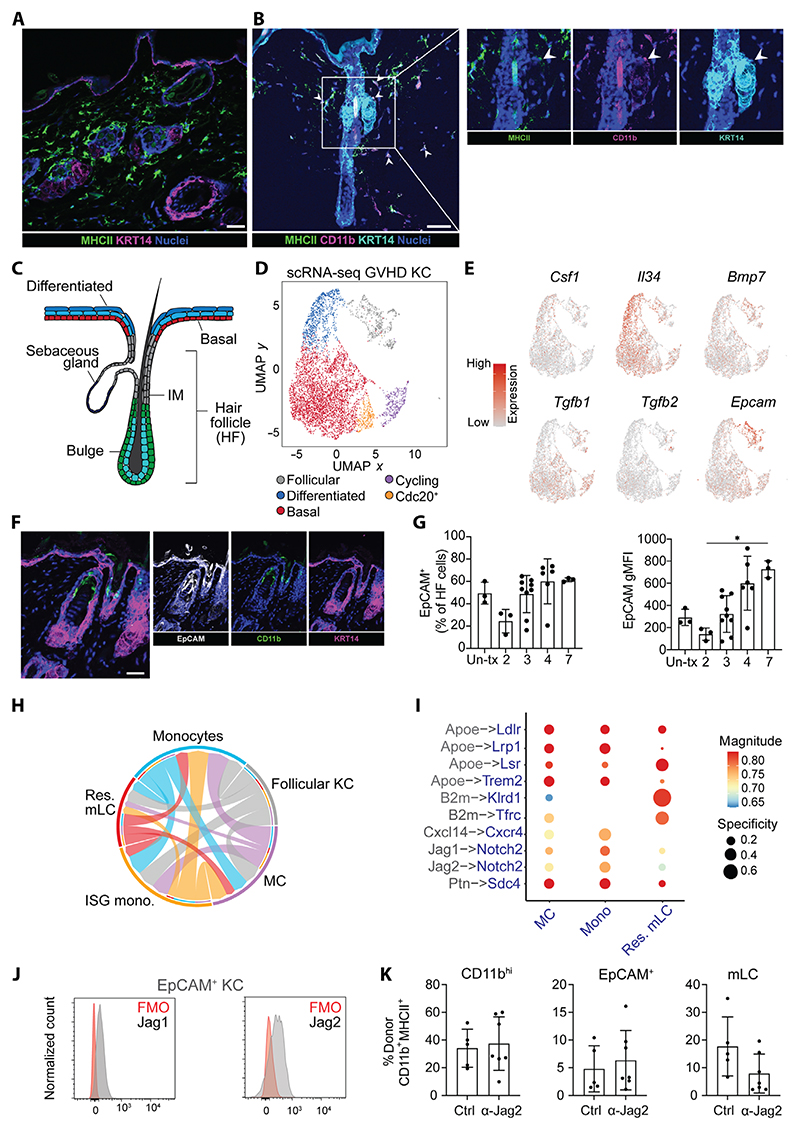
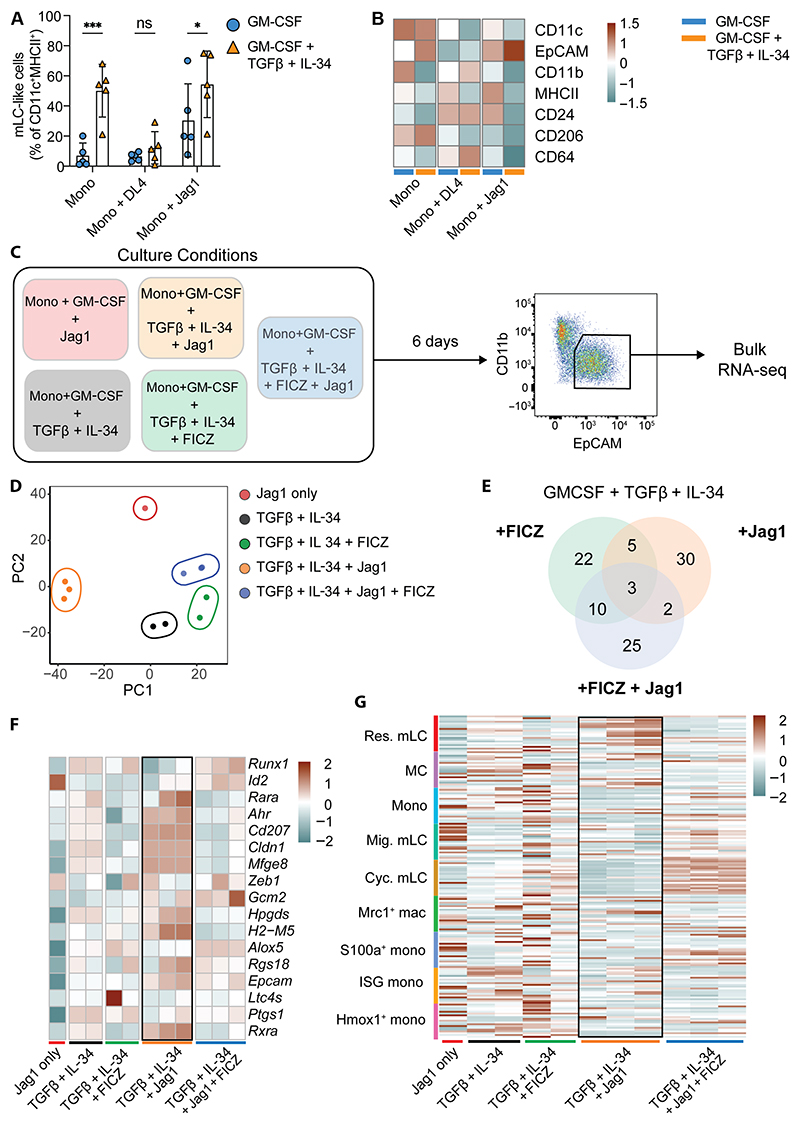
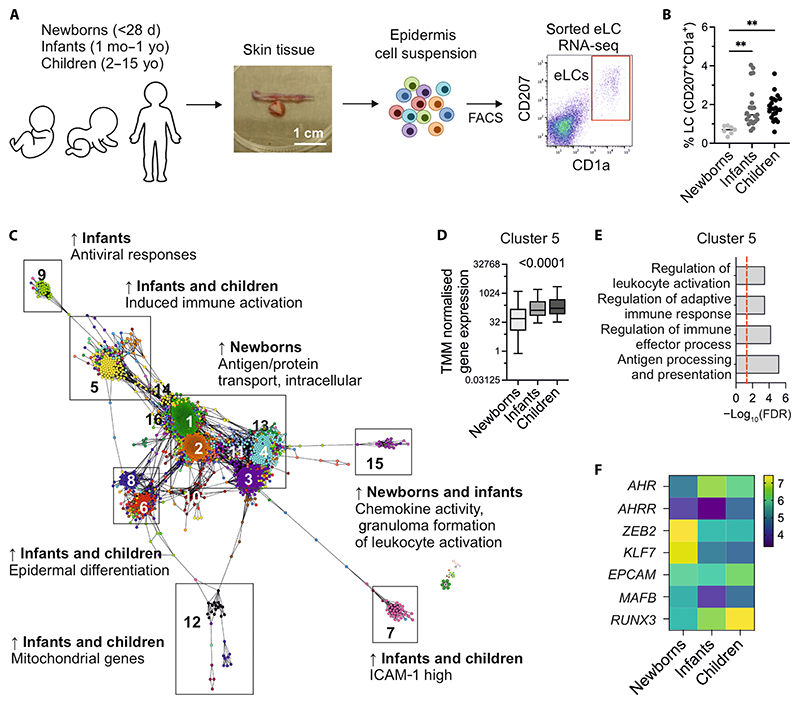
Langerhans cells (LCs) are distinct among phagocytes, functioning both as embryo-derived, tissue-resident macrophages in skin innervation and repair and as migrating professional antigen-presenting cells, a function classically assigned to dendritic cells (DCs). Here, we demonstrate that both intrinsic and extrinsic factors imprint this dual identity. Using ablation of embryo-derived LCs in the murine adult skin and tracking differentiation of incoming monocyte-derived replacements, we found intrinsic intraepidermal heterogeneity. We observed that ontogenically distinct monocytes give rise to LCs. Within the epidermis, Jagged-dependent activation of Notch signaling, likely within the hair follicle niche, provided an initial site of LC commitment before metabolic adaptation and survival of monocyte-derived LCs. In the human skin, embryo-derived LCs in newborns retained transcriptional evidence of their macrophage origin, but this was superseded by DC-like immune modules after postnatal expansion. Thus, adaptation to adult skin niches replicates conditioning of LC at birth, permitting repair of the embryo-derived LC network.
Conflict of interest statement
Figures
References
-
- Jakob T, Udey MC. Epidermal Langerhans cells: From neurons to nature’s adjuvants. Adv Dermatol. 1999;14:209–258. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
