

Excitable Rho dynamics control cell shape and motility by sequentially activating ERM proteins and actomyosin contractility
- PMID: 39241071
- PMCID: PMC11378911
- DOI: 10.1126/sciadv.adn6858
Excitable Rho dynamics control cell shape and motility by sequentially activating ERM proteins and actomyosin contractility
Abstract
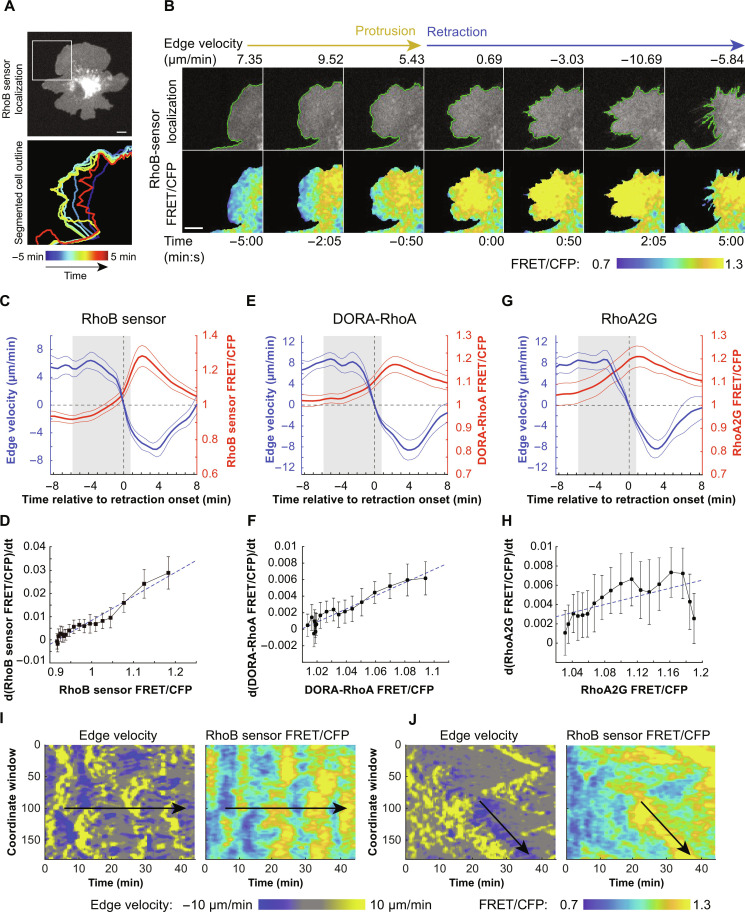
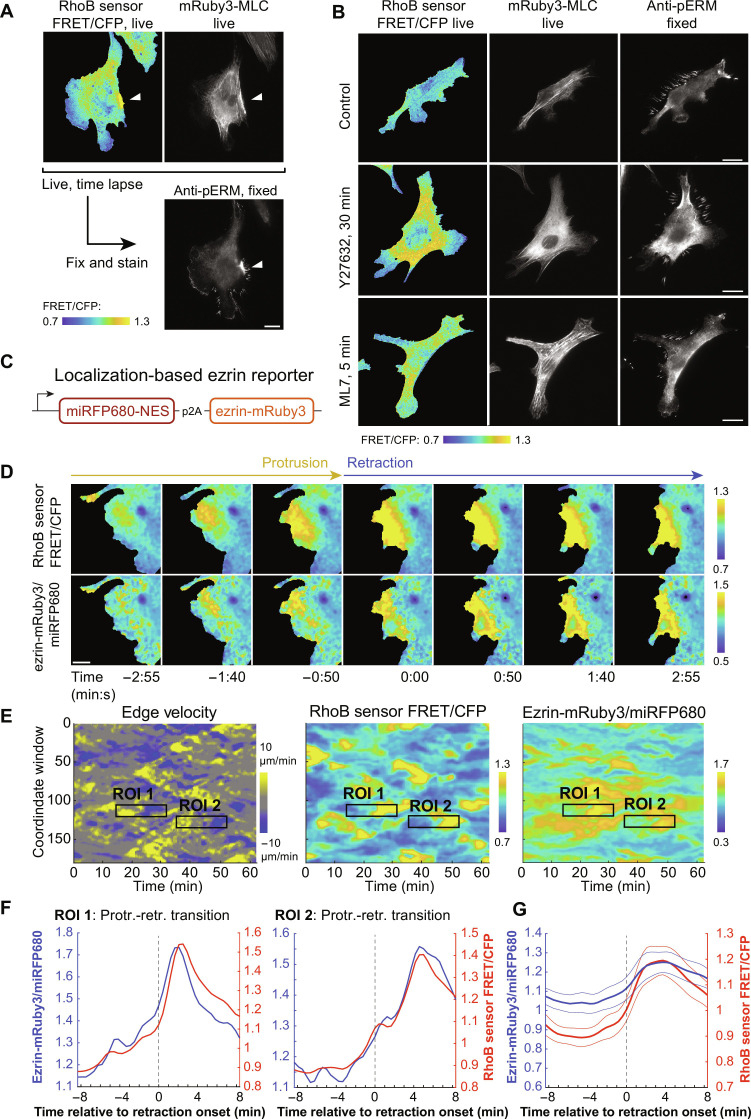
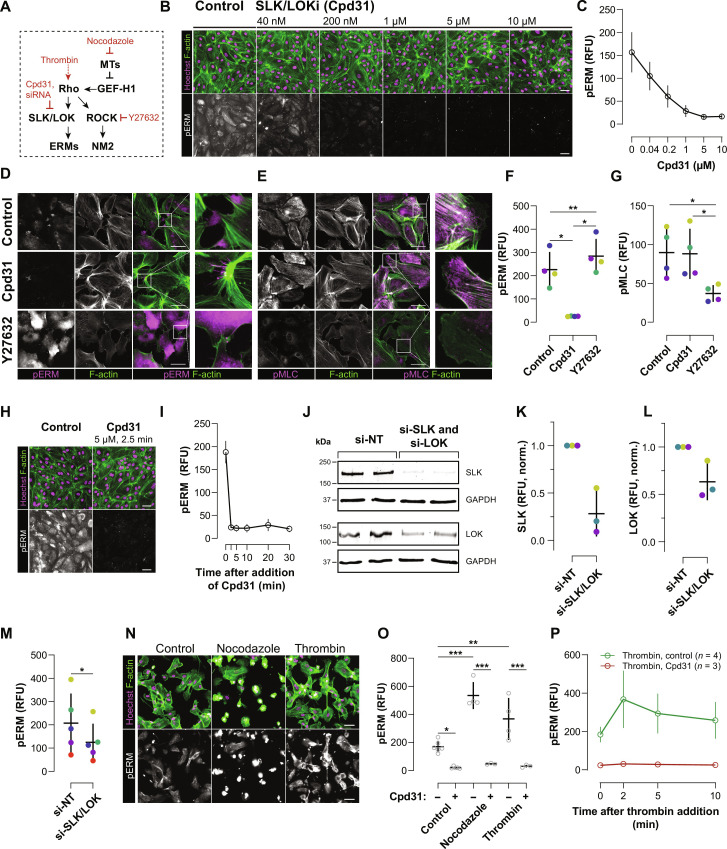
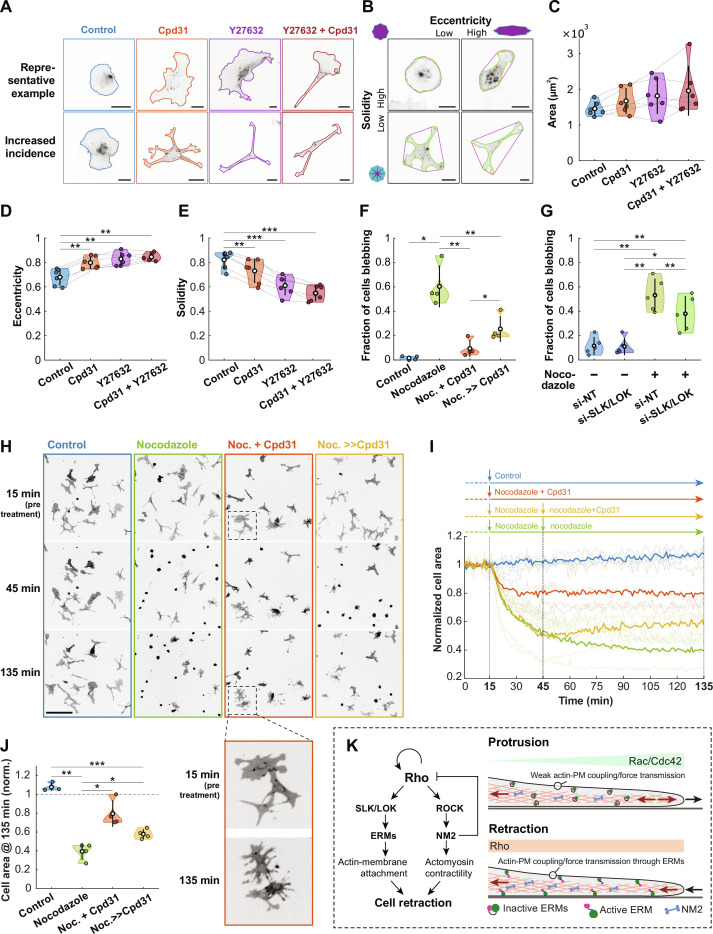
Migration of endothelial and many other cells requires spatiotemporal regulation of protrusive and contractile cytoskeletal rearrangements that drive local cell shape changes. Unexpectedly, the small GTPase Rho, a crucial regulator of cell movement, has been reported to be active in both local cell protrusions and retractions, raising the question of how Rho activity can coordinate cell migration. Here, we show that Rho activity is absent in local protrusions and active during retractions. During retractions, Rho rapidly activated ezrin-radixin-moesin proteins (ERMs) to increase actin-membrane attachment, and, with a delay, nonmuscle myosin 2 (NM2). Rho activity was excitable, with NM2 acting as a slow negative feedback regulator. Strikingly, inhibition of SLK/LOK kinases, through which Rho activates ERMs, caused elongated cell morphologies, impaired Rho-induced cell contractions, and reverted Rho-induced blebbing. Together, our study demonstrates that Rho activity drives retractions by sequentially enhancing ERM-mediated actin-membrane attachment for force transmission and NM2-dependent contractility.
Figures



References
-
- Verkhovsky A. B., The mechanisms of spatial and temporal patterning of cell-edge dynamics. Curr. Opin. Cell Biol. 36, 113–121 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
