

Human cell surface-AAV interactomes identify LRP6 as blood-brain barrier transcytosis receptor and immune cytokine IL3 as AAV9 binder
- PMID: 39245720
- PMCID: PMC11381518
- DOI: 10.1038/s41467-024-52149-0
Human cell surface-AAV interactomes identify LRP6 as blood-brain barrier transcytosis receptor and immune cytokine IL3 as AAV9 binder
Abstract
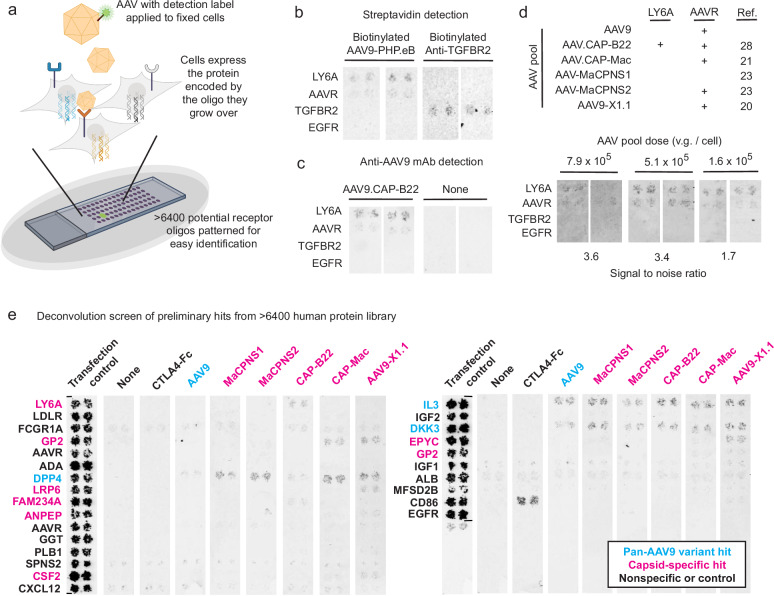
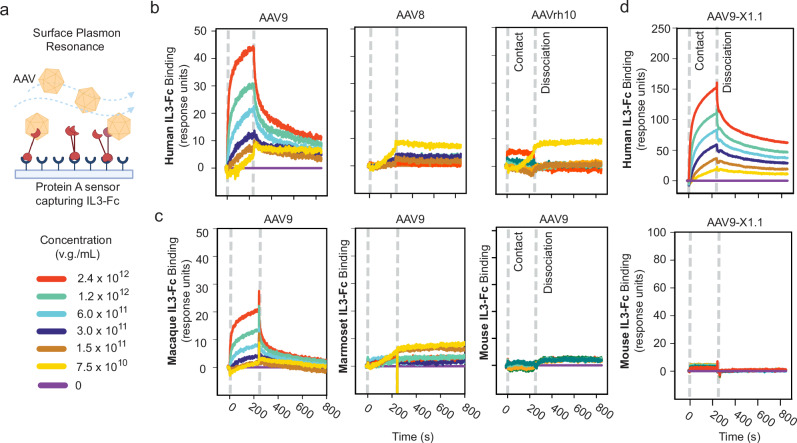
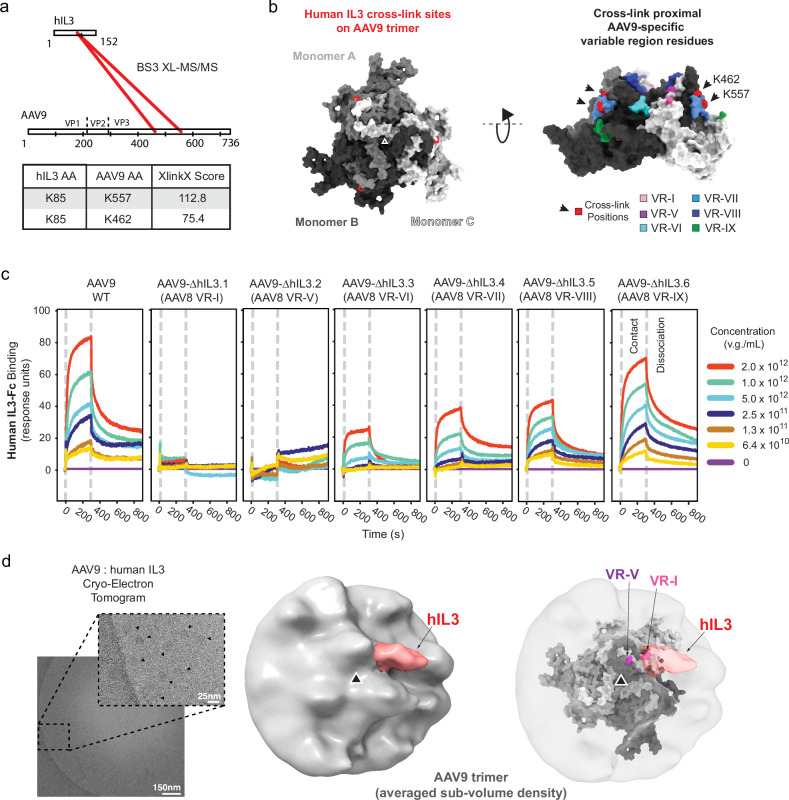
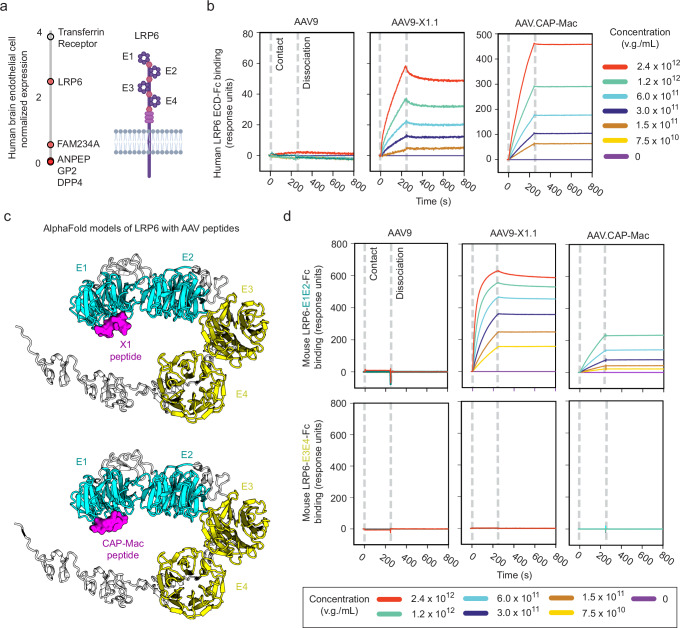
Adeno-associated viruses (AAVs) are foundational gene delivery tools for basic science and clinical therapeutics. However, lack of mechanistic insight, especially for engineered vectors created by directed evolution, can hamper their application. Here, we adapt an unbiased human cell microarray platform to determine the extracellular and cell surface interactomes of natural and engineered AAVs. We identify a naturally-evolved and serotype-specific interaction between the AAV9 capsid and human interleukin 3 (IL3), with possible roles in host immune modulation, as well as lab-evolved low-density lipoprotein receptor-related protein 6 (LRP6) interactions specific to engineered capsids with enhanced blood-brain barrier crossing in non-human primates after intravenous administration. The unbiased cell microarray screening approach also allows us to identify off-target tissue binding interactions of engineered brain-enriched AAV capsids that may inform vectors' peripheral organ tropism and side effects. Our cryo-electron tomography and AlphaFold modeling of capsid-interactor complexes reveal LRP6 and IL3 binding sites. These results allow confident application of engineered AAVs in diverse organisms and unlock future target-informed engineering of improved viral and non-viral vectors for non-invasive therapeutic delivery to the brain.
© 2024. The Author(s).
Conflict of interest statement
The California Institute of Technology has a patent pending for the delivery methods identified in this manuscript, with T.F.S, X.C., S.J., and V.G. listed as inventors (PCT Patent Application No: PCT/US2024/0139329) and a provisional patent for the sequences described in this manuscript, with S.J., T.J.B., T.F.S., and V.G listed as inventors. V.G. is a co-founder and board of directors member of Capsida Therapeutics, a fully integrated AAV engineering and gene therapy company. T.F.S and V.G. are co-founders and X.C. and X.D. are co-founders and employees of Receptive Biotherapeutics. B.W. and C.T. are employees of Charles River Laboratories. The remaining authors declare no competing interests.
Figures
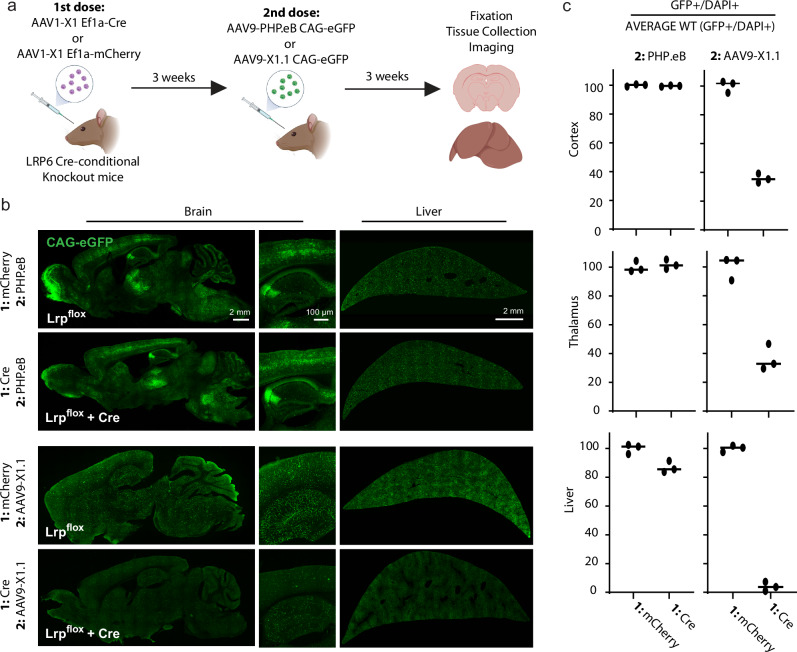
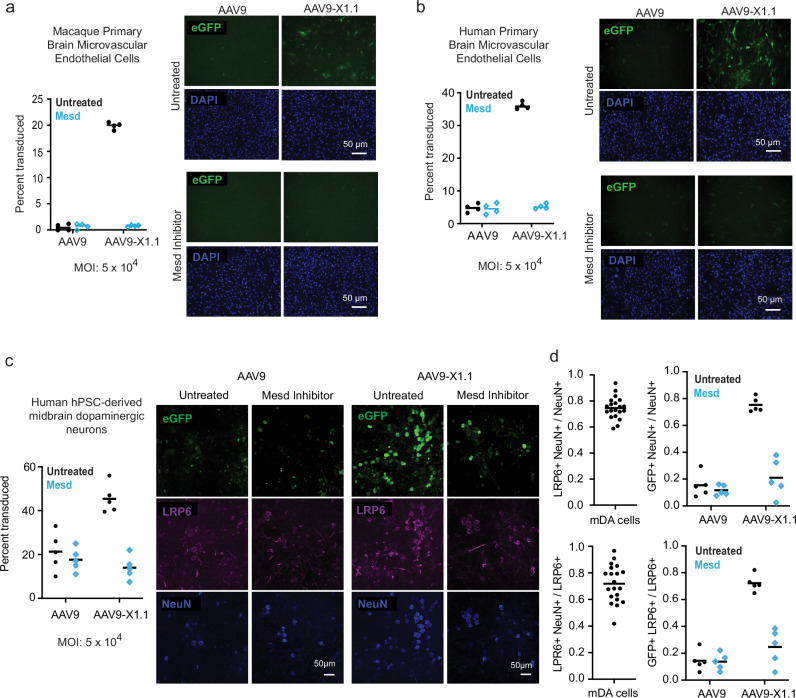
References
-
- Kuzmin, D. A. et al. The clinical landscape for AAV gene therapies. Nat. Rev. Drug Discov. d41573-021-00017–7 10.1038/d41573-021-00017-7 (2021). - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
- DP1 NS111369/NS/NINDS NIH HHS/United States
- UF1 MH128336/MH/NIMH NIH HHS/United States
- DP1NS111369/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- UF1MH128336/U.S. Department of Health & Human Services | NIH | National Institute of Mental Health (NIMH)
LinkOut - more resources
Full Text Sources
Miscellaneous
