

Improved health by combining dietary restriction and promoting muscle growth in DNA repair-deficient progeroid mice
- PMID: 39245994
- PMCID: PMC11634475
- DOI: 10.1002/jcsm.13570
Improved health by combining dietary restriction and promoting muscle growth in DNA repair-deficient progeroid mice
Abstract
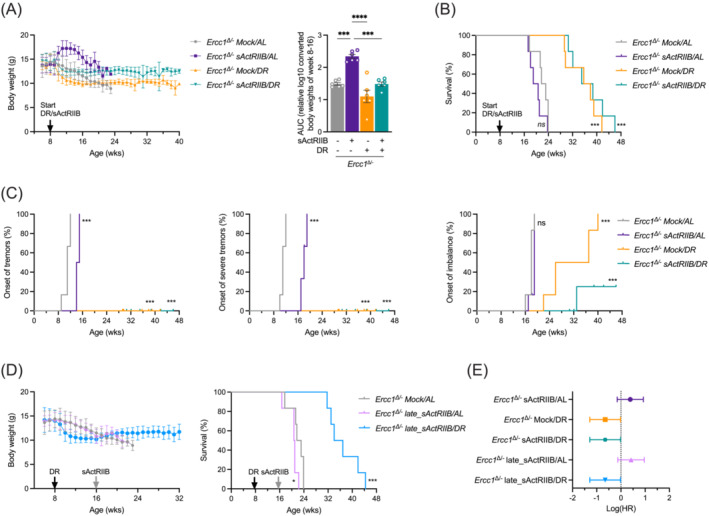
Background: Ageing is a complex multifactorial process, impacting all organs and tissues, with DNA damage accumulation serving as a common underlying cause. To decelerate ageing, various strategies have been applied to model organisms and evaluated for health and lifespan benefits. Dietary restriction (DR, also known as caloric restriction) is a well-established long-term intervention recognized for its universal anti-ageing effects. DR temporarily suppresses growth, and when applied to progeroid DNA repair-deficient mice doubles lifespan with systemic health benefits. Counterintuitively, attenuation of myostatin/activin signalling by soluble activin receptor (sActRIIB), boosts the growth of muscle and, in these animals, prevents muscle wasting, improves kidney functioning, and compresses morbidity.
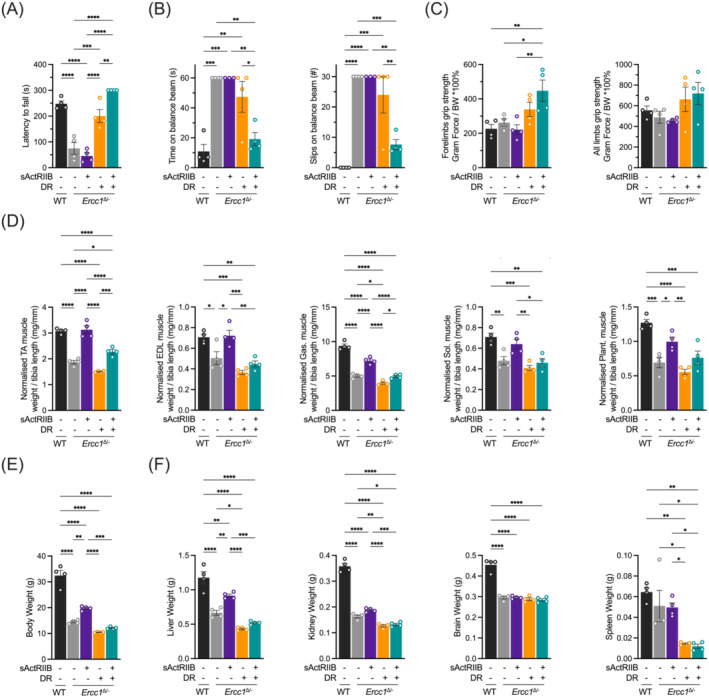
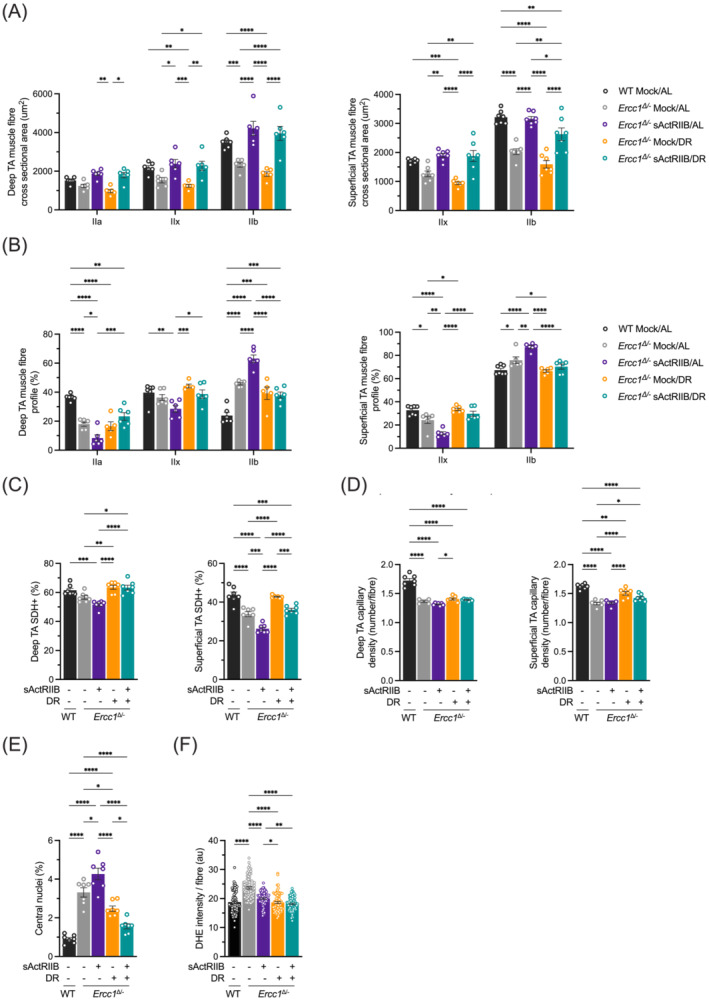
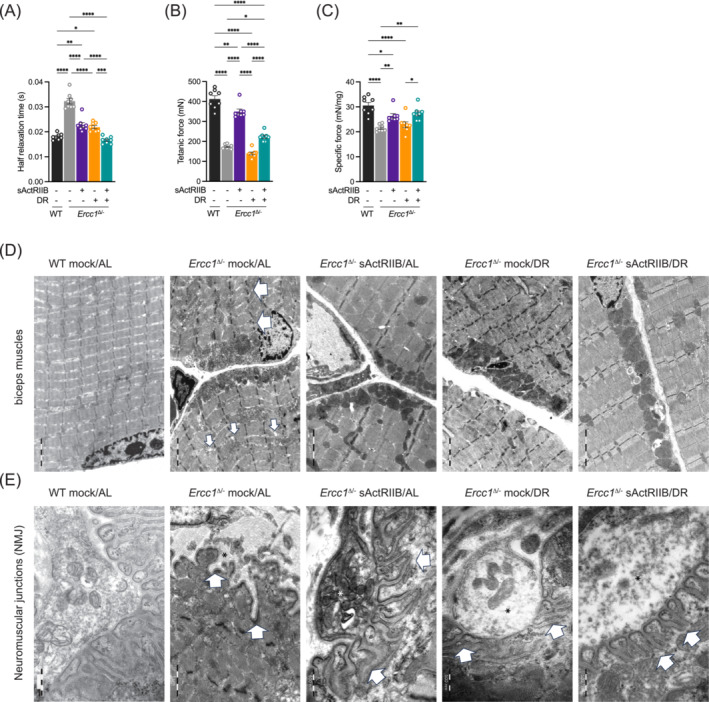
Methods: Here, we investigated a combined approach, applying an anabolic regime (sActRIIB) at the same time as DR to Ercc1Δ/- progeroid mice. Following both single treatments and combined, we monitored global effects on body weight, lifespan and behaviour, and local effects on muscle and tissue weight, muscle morphology and function, and ultrastructural and transcriptomic changes in muscle and kidney.
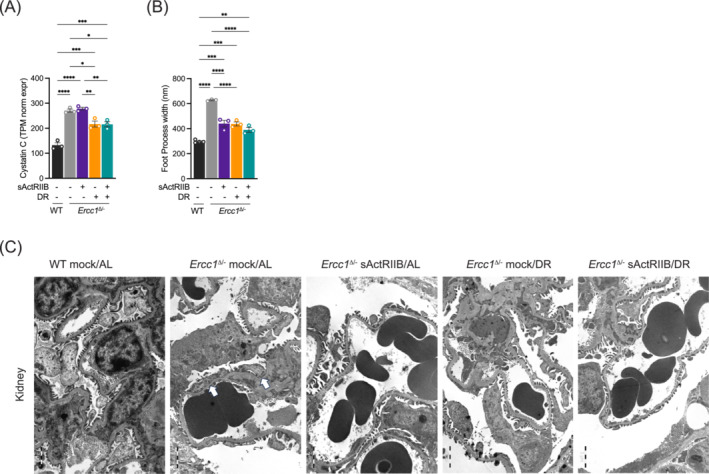
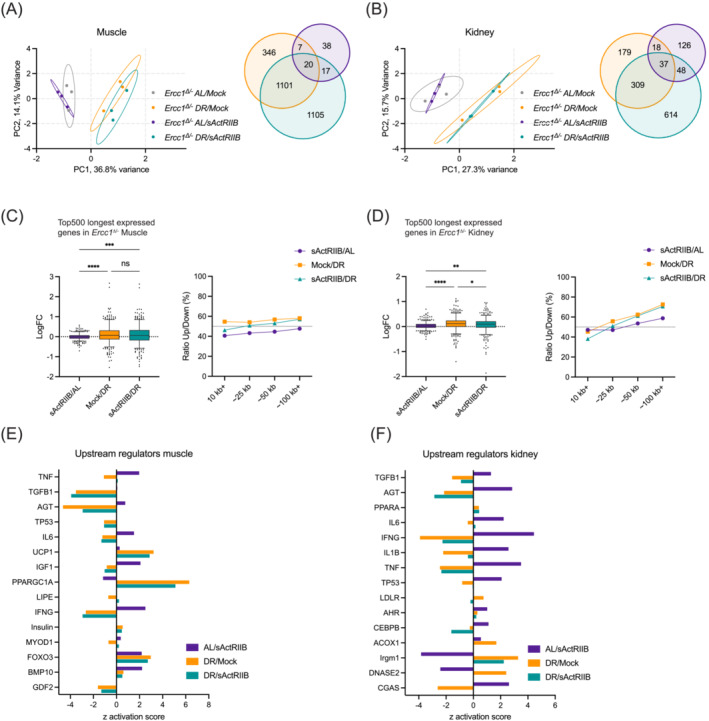
Results: Lifespan was mostly influenced by DR (extended from approximately 20 to 40 weeks; P < 0.001), with sActRIIB clearly increasing muscle mass (35-65%) and tetanic force (P < 0.001). The combined regime yielded a stable uniform body weight, but increased compared with DR alone, synergistically improved motor coordination and further delayed the onset and development of balance problems. sActRIIB significantly increased muscle fibre size (P < 0.05) in mice subjected to DR and lowered all signs of muscle damage. Ercc1Δ/- mice showed abnormal neuromuscular junctions. Single interventions by sActRIIB treatment or DR only partially rescued this phenotype, while in the double intervention group, the regularly shaped junctional foldings were maintained. In kidney of Ercc1Δ/- mice, we observed a mild but significant foot process effacement, which was restored by either intervention. Transcriptome analysis also pointed towards reduced levels of DNA damage in muscle and kidney by DR, but not sActRIIB, while these levels retained lower in the double intervention.
Conclusions: In muscle, we found synergistic effects of combining sActRIIB with DR, but not in kidney, with an overall better health in the double intervention group. Crucially, the benefits of each single intervention are not lost when administered in combination, but rather strengthened, even when sActRIIB was applied late in life, opening opportunities for translation to human.
Keywords: Ageing; Dietary restriction; Kidney; Muscle; Myostatin; Progeria.
© 2024 The Author(s). Journal of Cachexia, Sarcopenia and Muscle published by Wiley Periodicals LLC.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- KR1984/4-1/Deutsche Forschungsgemeinschaft
- HU1016/8-2/Deutsche Forschungsgemeinschaft
- CRC1192/DFG
- HU1016/11-1/DFG
- HU1016/12-1/DFG
- STOP-FSGS-01GM2202A/BMBF
- NephrESA-031L0191E/BMBF
- UPTAKE-01EK2105D/BMBF
- ONCODE by the Dutch Cancer Society
- 737.016.015/Netherlands Organisation for Scientific Research (NWO)
- 733050810/Memorabel (ZonMW)
- ADPS Longevity Research Award
- 162135/Regiodeal Foodvalley
- TC-NERRD20-113/European Joint Programme Rare Diseases (EJPRD)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
