

This is a preprint.
CO2 protects cells from iron-Fenton oxidative DNA damage in E. coli and humans
- PMID: 39253463
- PMCID: PMC11383276
- DOI: 10.1101/2024.08.26.609766
CO2 protects cells from iron-Fenton oxidative DNA damage in E. coli and humans
Update in
-
CO2 protects cells from iron-Fenton oxidative DNA damage in Escherichia coli and humans.Proc Natl Acad Sci U S A. 2024 Dec 3;121(49):e2419175121. doi: 10.1073/pnas.2419175121. Epub 2024 Nov 27. Proc Natl Acad Sci U S A. 2024. PMID: 39602264 Free PMC article.
Abstract
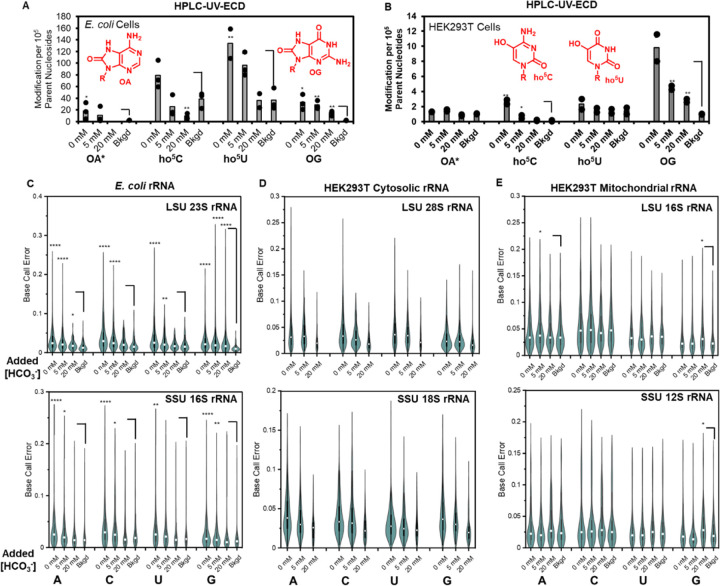
Whereas hydroxyl radical is commonly named as the Fenton product responsible for DNA and RNA damage in cells, here we demonstrate that the cellular reaction generates carbonate radical anion due to physiological levels of bicarbonate. Analysis of the metabolome, transcriptome and, in human cells, the nuclear genome shows a consistent buffering of H2O2-induced oxidative stress leading to one common pathway, namely guanine oxidation. Particularly revealing are nanopore-based studies of direct RNA sequencing of cytosolic and mitochondrial ribosomal RNA along with glycosylase-dependent qPCR studies of oxidative DNA damage in telomeres. The focusing of oxidative modification on one pathway is consistent with the highly evolved base excision repair suite of enzymes and their involvement in gene regulation in response to oxidative stress.
Conflict of interest statement
Competing Interests. The authors declare no competing interests in this work.
Figures




References
-
- Fenton H. J. H., LXXIII.—Oxidation of tartaric acid in presence of iron. J. Chem. Soc., Trans. 65, 899–910 (1894).
-
- Bataineh H., Pestovsky O., Bakac A., pH-induced mechanistic changeover from hydroxyl radicals to iron(iv) in the Fenton reaction. Chem. Soc. 3, 1594–1599 (2012).
-
- Kakhlon O., Cabantchik Z. I., The labile iron pool: characterization, measurement, and participation in cellular processes(1). Free Radic. Biol. Med. 33, 1037–1046 (2002). - PubMed
-
- Giorgio M., Trinei M., Migliaccio E., Pelicci P. G., Hydrogen peroxide: a metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 8, 722–728 (2007). - PubMed
-
- Roos W. P., Thomas A. D., Kaina B., DNA damage and the balance between survival and death in cancer biology. Nat. Rev. Cancer 16, 20–33 (2016). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources