

Glioblastoma induces the recruitment and differentiation of dendritic-like "hybrid" neutrophils from skull bone marrow
- PMID: 39255776
- PMCID: PMC11446475
- DOI: 10.1016/j.ccell.2024.08.008
Glioblastoma induces the recruitment and differentiation of dendritic-like "hybrid" neutrophils from skull bone marrow
Abstract
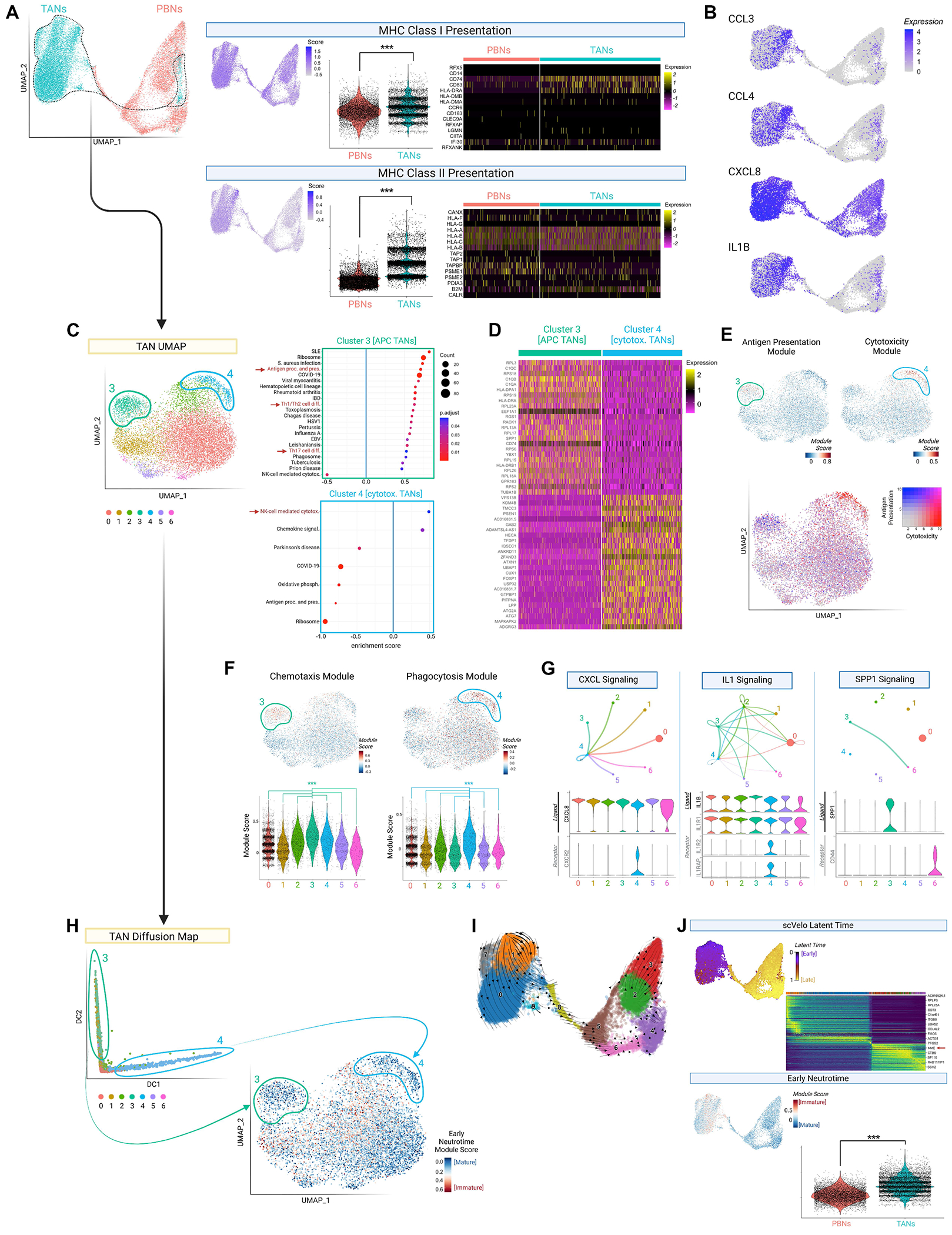
Tumor-associated neutrophil (TAN) effects on glioblastoma (GBM) biology remain under-characterized. We show here that neutrophils with dendritic features-including morphological complexity, expression of antigen presentation genes, and the ability to process exogenous peptide and stimulate major histocompatibility complex (MHC)II-dependent T cell activation-accumulate intratumorally and suppress tumor growth in vivo. Trajectory analysis of patient TAN scRNA-seq identifies this "hybrid" dendritic-neutrophil phenotype as a polarization state that is distinct from canonical cytotoxic TANs, and which differentiates from local precursors. These hybrid-inducible immature neutrophils-which we identified in patient and murine glioblastomas-arise not from circulation, but from local skull marrow. Through labeled skull flap transplantation and targeted ablation, we characterize calvarial marrow as a contributor of antitumoral myeloid antigen-presenting cells (APCs), including TANs, which elicit T cell cytotoxicity and memory. As such, agents augmenting neutrophil egress from skull marrow-such as intracalvarial AMD3100, whose survival-prolonging effect in GBM we report-present therapeutic potential.
Keywords: MHC class II; T cells; antigen-presenting cells; dendritic cells; glioblastoma; myeloid; skull marrow; tumor-associated neutrophil.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






Update of
-
Glioblastoma induces the recruitment and differentiation of hybrid neutrophils from skull bone marrow.bioRxiv [Preprint]. 2023 Mar 25:2023.03.24.534105. doi: 10.1101/2023.03.24.534105. bioRxiv. 2023. Update in: Cancer Cell. 2024 Sep 9;42(9):1549-1569.e16. doi: 10.1016/j.ccell.2024.08.008. PMID: 36993266 Free PMC article. Updated. Preprint.
References
-
- Luo JW, Wang X, Yang Y, and Mao Q (2015). Role of micro-RNA (miRNA) in pathogenesis of glioblastoma. European review for medical and pharmacological sciences 19, 1630–1639. - PubMed
-
- Raychaudhuri B, Rayman P, Huang P, Grabowski M, Hambardzumyan D, Finke JH, and Vogelbaum MA (2015). Myeloid derived suppressor cell infiltration of murine and human gliomas is associated with reduction of tumor infiltrating lymphocytes. J Neurooncol 122, 293–301. 10.1007/s11060-015-1720-6. - DOI - PubMed
-
- Singhal S, Bhojnagarwala PS, O’Brien S, Moon EK, Garfall AL, Rao AS, Quatromoni JG, Stephen TL, Litzky L, Deshpande C, et al. (2016). Origin and Role of a Subset of Tumor-Associated Neutrophils with Antigen-Presenting Cell Features in Early-Stage Human Lung Cancer. Cancer Cell 30, 120–135. 10.1016/j.ccell.2016.06.001. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
