

A whole-body micro-CT scan library that captures the skeletal diversity of Lake Malawi cichlid fishes
- PMID: 39256465
- PMCID: PMC11387623
- DOI: 10.1038/s41597-024-03687-1
A whole-body micro-CT scan library that captures the skeletal diversity of Lake Malawi cichlid fishes
Abstract
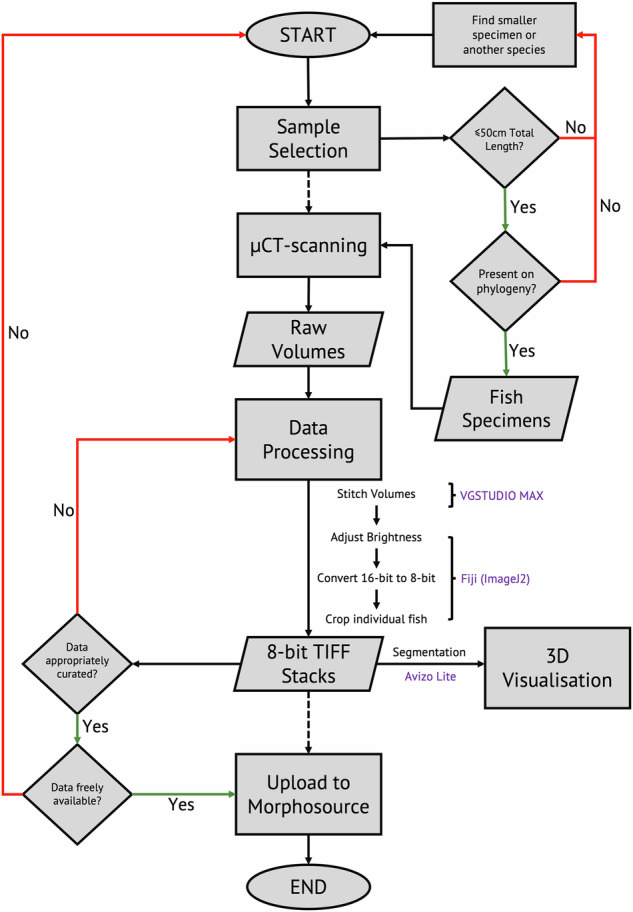
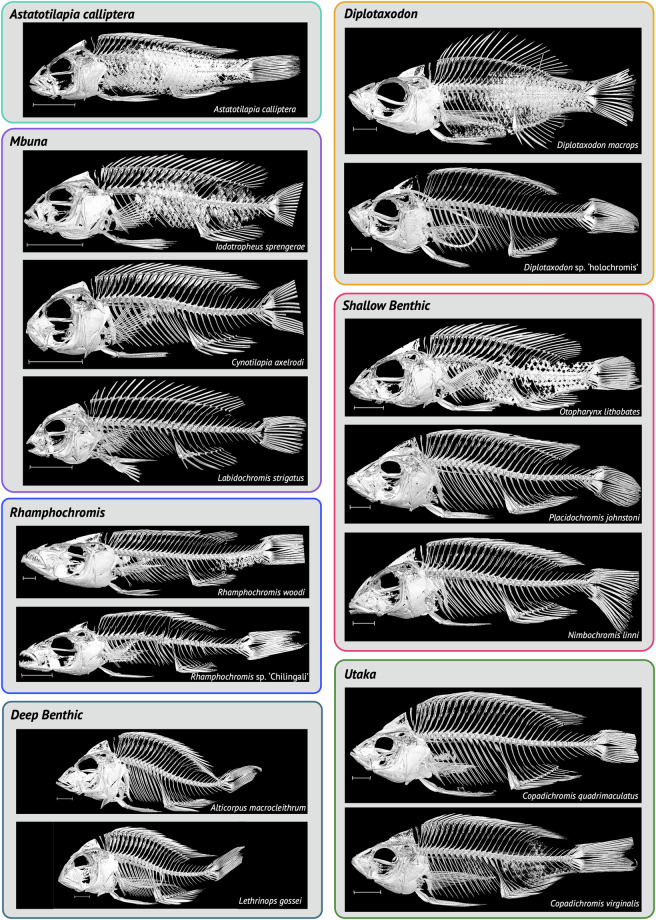
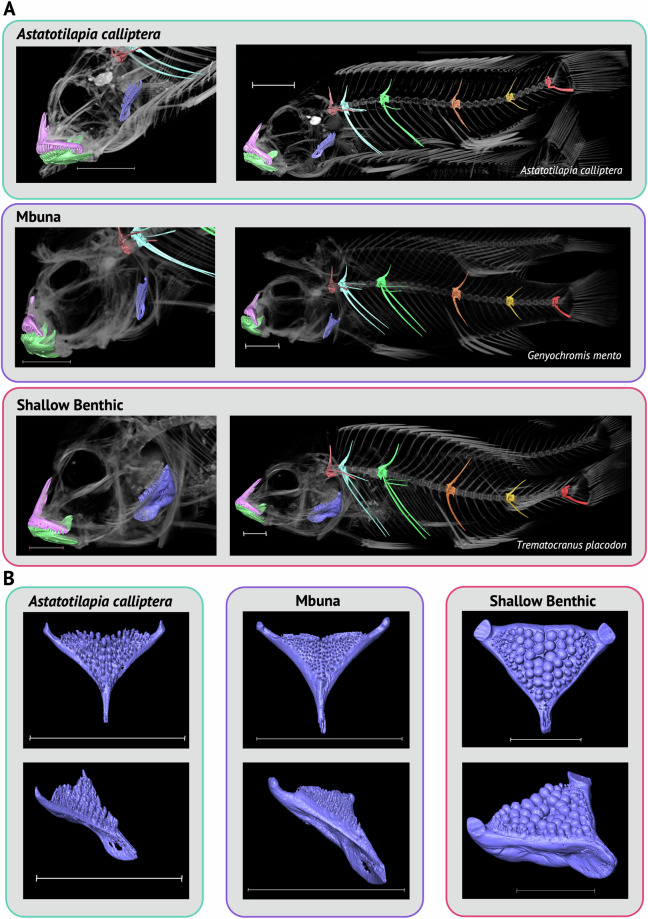
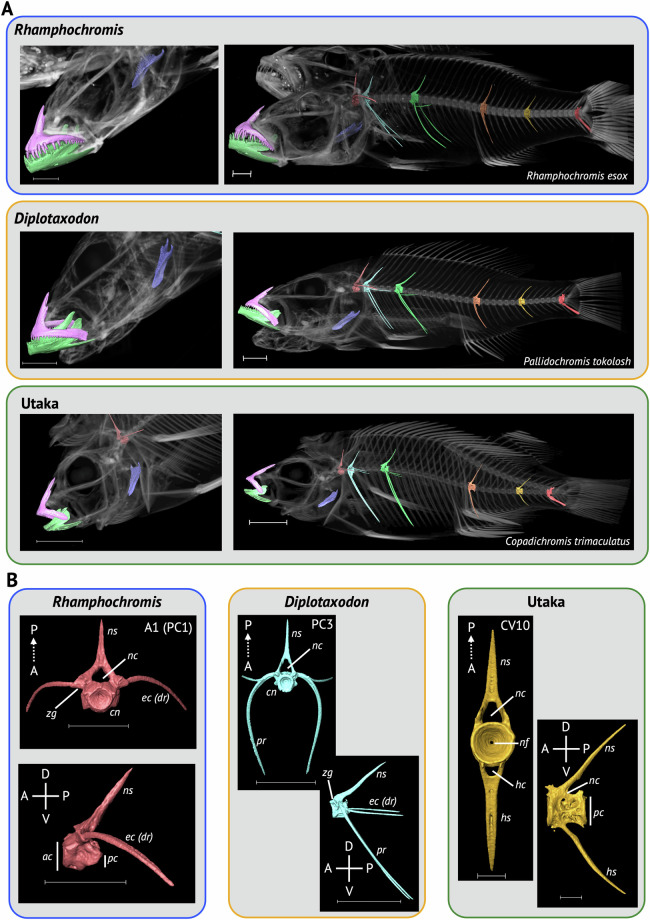
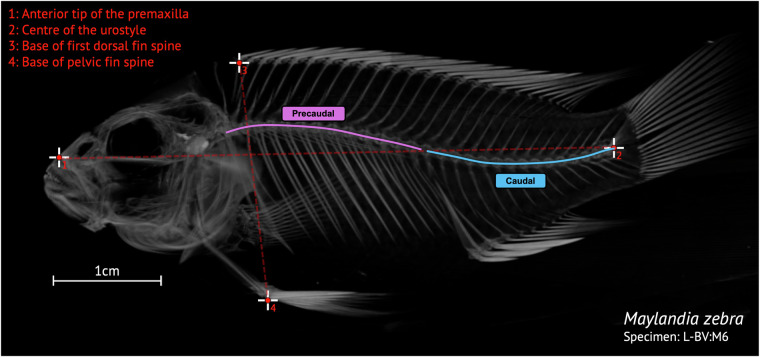
Here we describe a dataset of freely available, readily processed, whole-body μCT-scans of 56 species (116 specimens) of Lake Malawi cichlid fishes that captures a considerable majority of the morphological variation present in this remarkable adaptive radiation. We contextualise the scanned specimens within a discussion of their respective ecomorphological groupings and suggest possible macroevolutionary studies that could be conducted with these data. In addition, we describe a methodology to efficiently μCT-scan (on average) 23 specimens per hour, limiting scanning time and alleviating the financial cost whilst maintaining high resolution. We demonstrate the utility of this method by reconstructing 3D models of multiple bones from multiple specimens within the dataset. We hope this dataset will enable further morphological study of this fascinating system and permit wider-scale comparisons with other cichlid adaptive radiations.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Barlow, G. The Cichlid Fishes: Nature’s Grand Experiment In Evolution (Hachette UK, London, 2008).
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
