

PKM2 regulates metabolic flux and oxidative stress in the murine heart
- PMID: 39256891
- PMCID: PMC11387154
- DOI: 10.14814/phy2.70040
PKM2 regulates metabolic flux and oxidative stress in the murine heart
Erratum in
-
Correction to "PKM2 regulates metabolic flux and oxidative stress in the murine heart".Physiol Rep. 2024 Oct;12(20):e70040. doi: 10.14814/phy2.70100. Physiol Rep. 2024. PMID: 39448389 Free PMC article. No abstract available.
Abstract
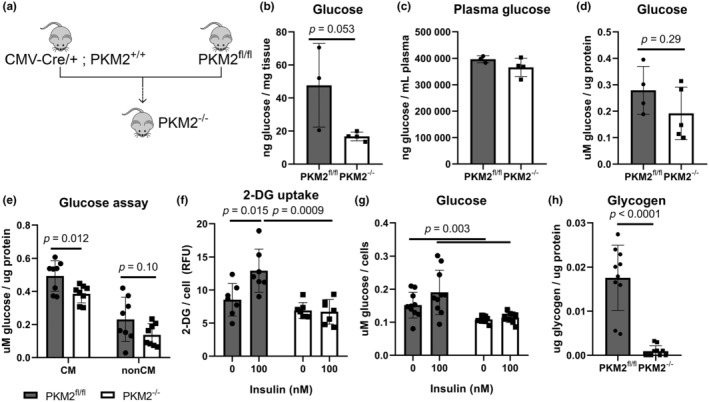
Cardiac metabolism ensures a continuous ATP supply, primarily using fatty acids in a healthy state and favoring glucose in pathological conditions. Pyruvate kinase muscle (PKM) controls the final step of glycolysis, with PKM1 being the main isoform in the heart. PKM2, elevated in various heart diseases, has been suggested to play a protective role in cardiac stress, but its function in basal cardiac metabolism remains unclear. We examined hearts from global PKM2 knockout (PKM2-/-) mice and found reduced intracellular glucose. Isotopic tracing of U-13C glucose revealed a shift to biosynthetic pathways in PKM2-/- cardiomyocytes. Total ATP content was two-thirds lower in PKM2-/- hearts, and functional analysis indicated reduced mitochondrial oxygen consumption. Total reactive oxygen species (ROS) and mitochondrial superoxide were also increased in PKM2-/- cardiomyocytes. Intriguingly, PKM2-/- hearts had preserved ejection fraction compared to controls. Mechanistically, increased calcium/calmodulin-dependent kinase II activity and phospholamban phosphorylation may contribute to higher sarcoendoplasmic reticulum calcium ATPase 2 pump activity in PKM2-/- hearts. Loss of PKM2 led to altered glucose metabolism, diminished mitochondrial function, and increased ROS in cardiomyocytes. These data suggest that cardiac PKM2 acts as an important rheostat to maintain ATP levels while limiting oxidative stress. Although loss of PKM2 did not impair baseline contractility, its absence may make hearts more sensitive to environmental stress or injury.
Keywords: glucose; glycolysis; metabolism; reactive oxygen species.
© 2024 The Author(s). Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures






References
-
- Bednarski, T. , Pyrkowska, A. , Opasińska, A. , & Dobrzyń, P. (2016). Regulation of cardiac metabolism and function by lipogenic factors. Advances in Hygiene and Experimental Medicine, 70, 644–653. - PubMed
-
- Boudina, S. , & Abel, E. D. (2007). Diabetic cardiomyopathy revisited. Circulation, 115, 3213–3223. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
