

This is a preprint.
Prodrug activation in malaria parasites mediated by an imported erythrocyte esterase, acylpeptide hydrolase (APEH)
- PMID: 39257815
- PMCID: PMC11383709
- DOI: 10.1101/2024.08.30.610542
Prodrug activation in malaria parasites mediated by an imported erythrocyte esterase, acylpeptide hydrolase (APEH)
Update in
-
Prodrug activation in malaria parasites mediated by an imported erythrocyte esterase, acylpeptide hydrolase (APEH).Proc Natl Acad Sci U S A. 2025 Mar 11;122(10):e2417682122. doi: 10.1073/pnas.2417682122. Epub 2025 Mar 4. Proc Natl Acad Sci U S A. 2025. PMID: 40035755 Free PMC article.
Abstract
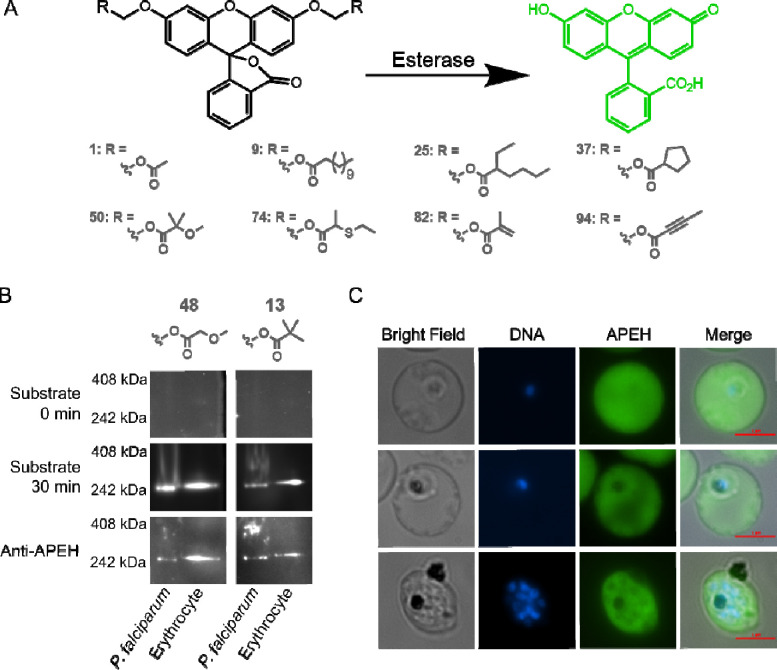
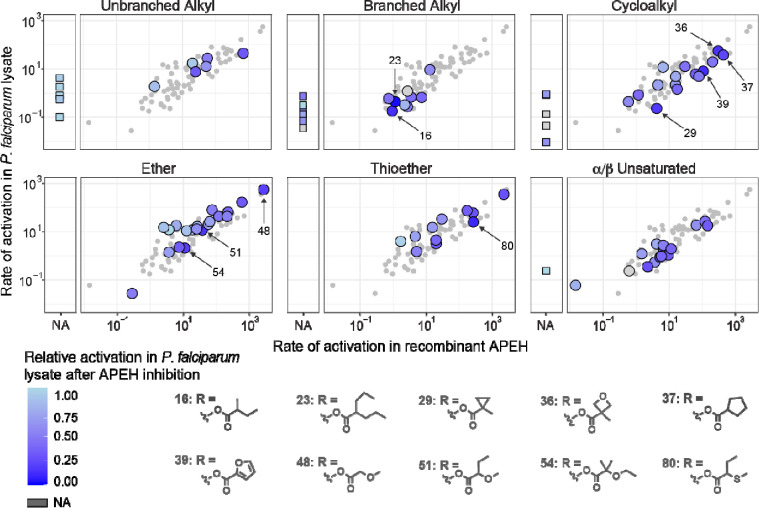
The continued emergence of antimalarial drug resistance highlights the need to develop new antimalarial therapies. Unfortunately, new drug development is often hampered by poor drug-like properties of lead compounds. Prodrugging temporarily masks undesirable compound features, improving bioavailability and target penetration. We have found that lipophilic diester prodrugs of phosphonic acid antibiotics, such as fosmidomycin, exhibit significantly higher antimalarial potency than their parent compounds (1). However, the activating enzymes for these prodrugs were unknown. Here, we show that an erythrocyte enzyme, acylpeptide hydrolase (APEH) is the major activating enzyme of multiple lipophilic ester prodrugs. Surprisingly, this enzyme is taken up by the malaria parasite, Plasmodium falciparum, where it localizes to the parasite cytoplasm and retains enzymatic activity. Using a novel fluorogenic ester library, we characterize the structure activity relationship of APEH, and compare it to that of P. falciparum esterases. We show that parasite-internalized APEH plays an important role in the activation of substrates with branching at the alpha carbon, in keeping with its exopeptidase activity. Our findings highlight a novel mechanism for antimicrobial prodrug activation, relying on a host-derived enzyme to yield activation at a microbial target. Mutations in prodrug activating enzymes are a common mechanism for antimicrobial drug resistance (2-4). Leveraging an internalized host enzyme would circumvent this, enabling the design of prodrugs with higher barriers to drug resistance.
Figures


References
-
- World Malaria Report 2023 (World Health Organization, 2023).
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous