

This is a preprint.
Mitochondrial RNA cytosolic leakage drives the SASP
- PMID: 39257994
- PMCID: PMC11384804
- DOI: 10.21203/rs.3.rs-4876596/v1
Mitochondrial RNA cytosolic leakage drives the SASP
Update in
-
Mitochondrial RNA cytosolic leakage drives the SASP.Nat Commun. 2025 Dec 15;16(1):10992. doi: 10.1038/s41467-025-66159-z. Nat Commun. 2025. PMID: 41398033 Free PMC article.
Abstract
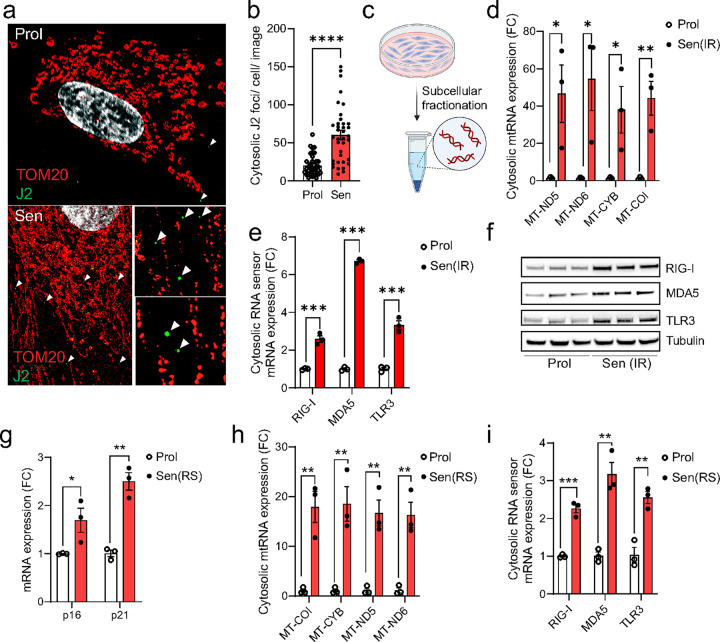
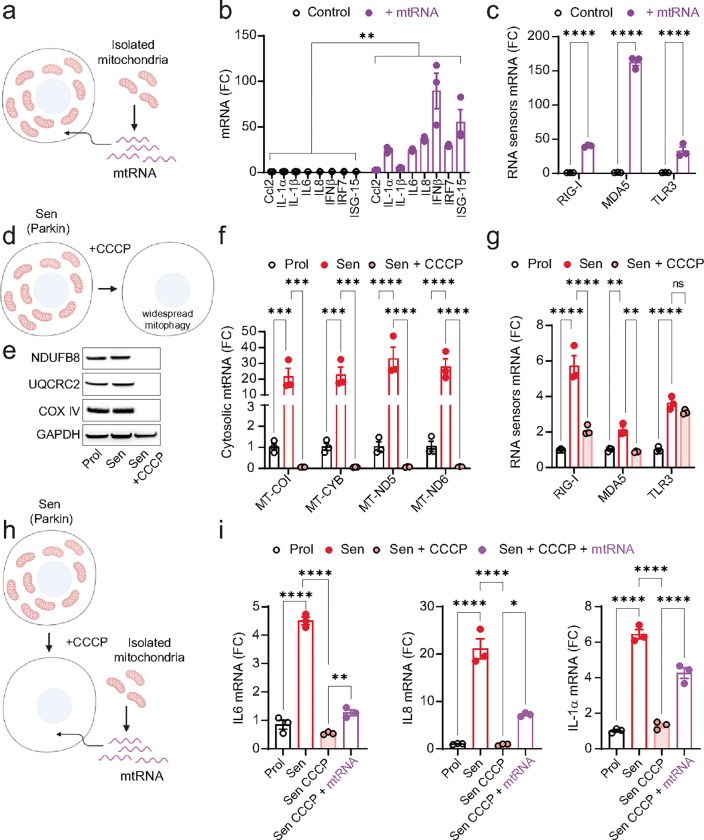
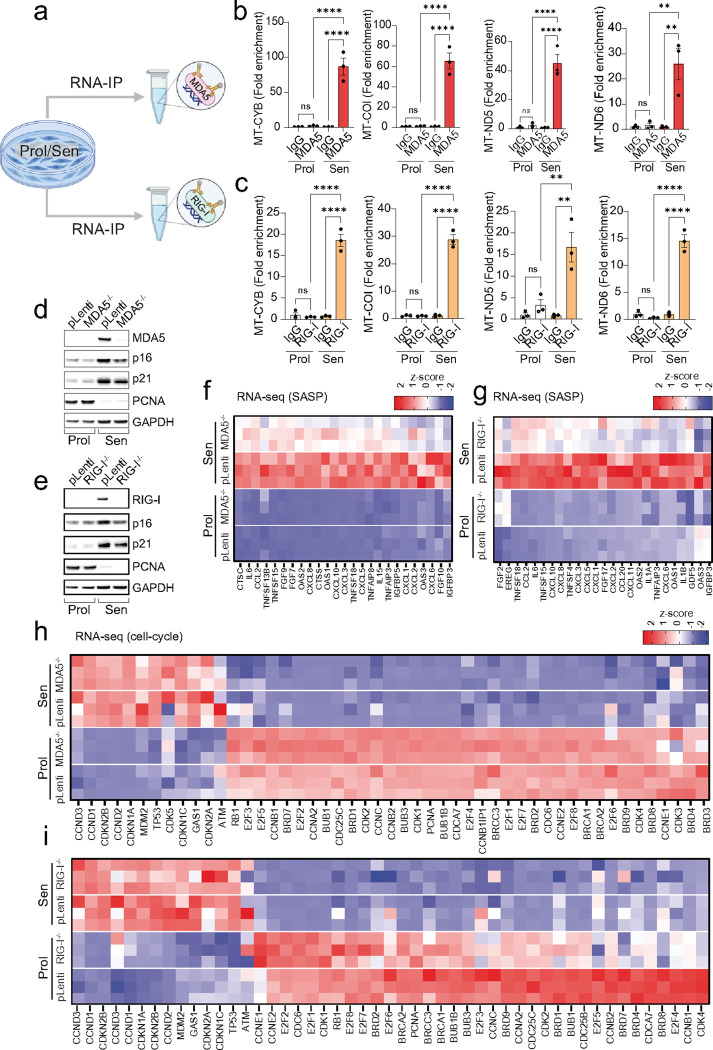
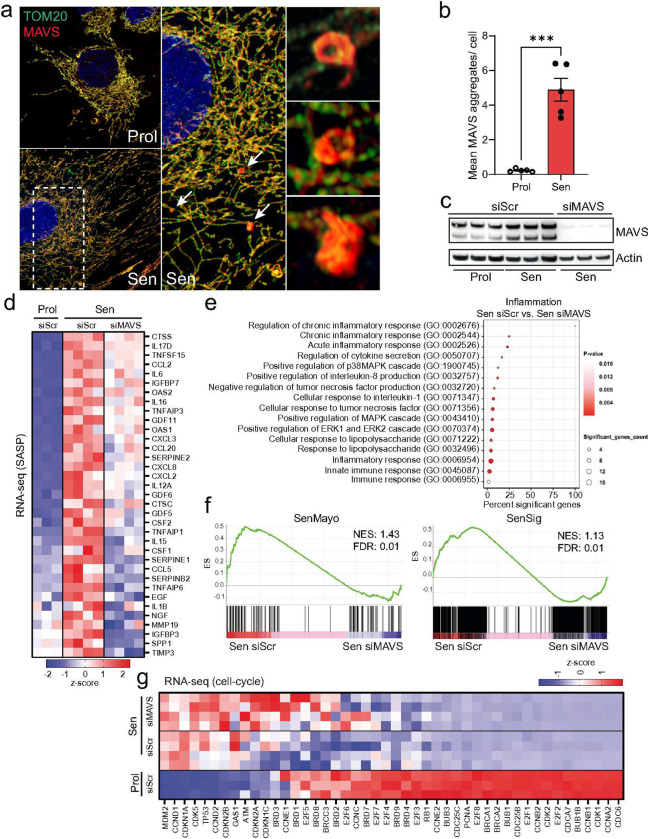
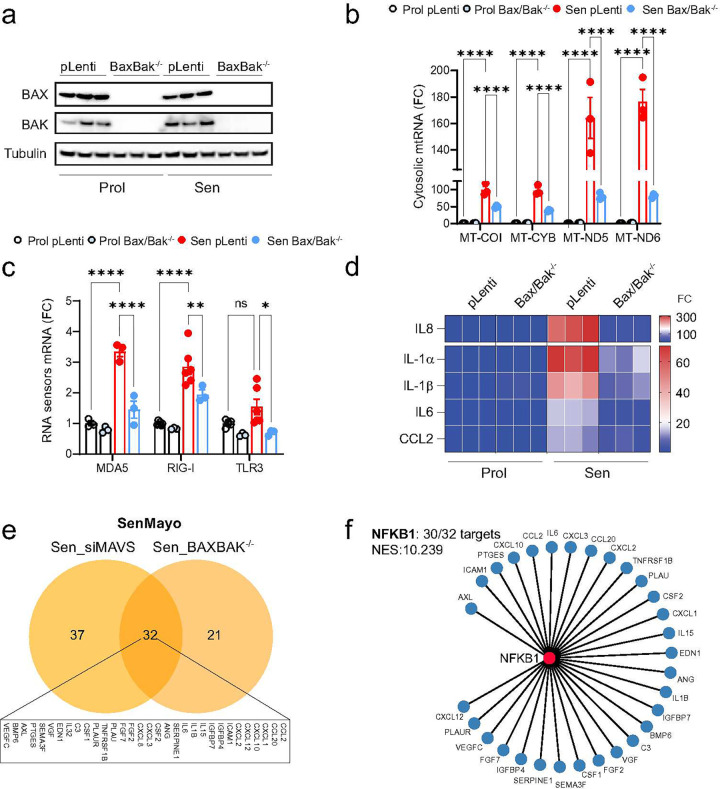
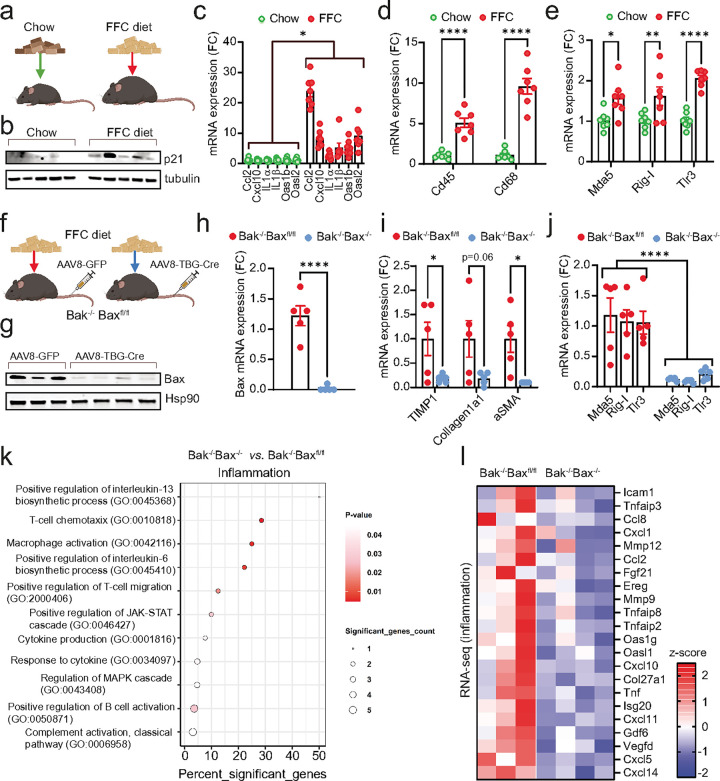
Senescent cells secrete proinflammatory factors known as the senescence-associated secretory phenotype (SASP), contributing to tissue dysfunction and aging. Mitochondrial dysfunction is a key feature of senescence, influencing SASP via mitochondrial DNA (mtDNA) release and cGAS/STING pathway activation. Here, we demonstrate that mitochondrial RNA (mtRNA) also accumulates in the cytosol of senescent cells, activating RNA sensors RIG-I and MDA5, leading to MAVS aggregation and SASP induction. Inhibition of these RNA sensors significantly reduces SASP factors. Furthermore, BAX and BAK plays a key role in mtRNA leakage during senescence, and their deletion diminishes SASP expression in vitro and in a mouse model of Metabolic Dysfunction Associated Steatohepatitis (MASH). These findings highlight mtRNA's role in SASP regulation and its potential as a therapeutic target for mitigating age-related inflammation.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
