

TNFSF14+ natural killer cells prevent spontaneous abortion by restricting leucine-mediated decidual stromal cell senescence
- PMID: 39261664
- PMCID: PMC11535022
- DOI: 10.1038/s44318-024-00220-3
TNFSF14+ natural killer cells prevent spontaneous abortion by restricting leucine-mediated decidual stromal cell senescence
Abstract
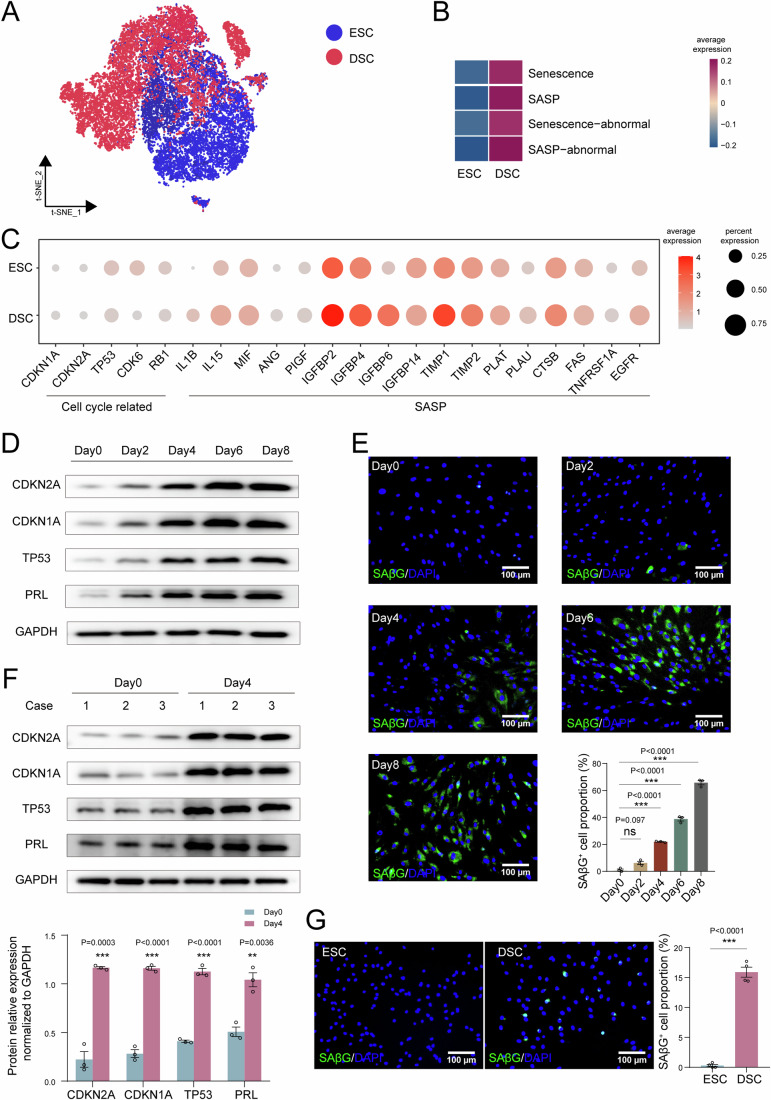
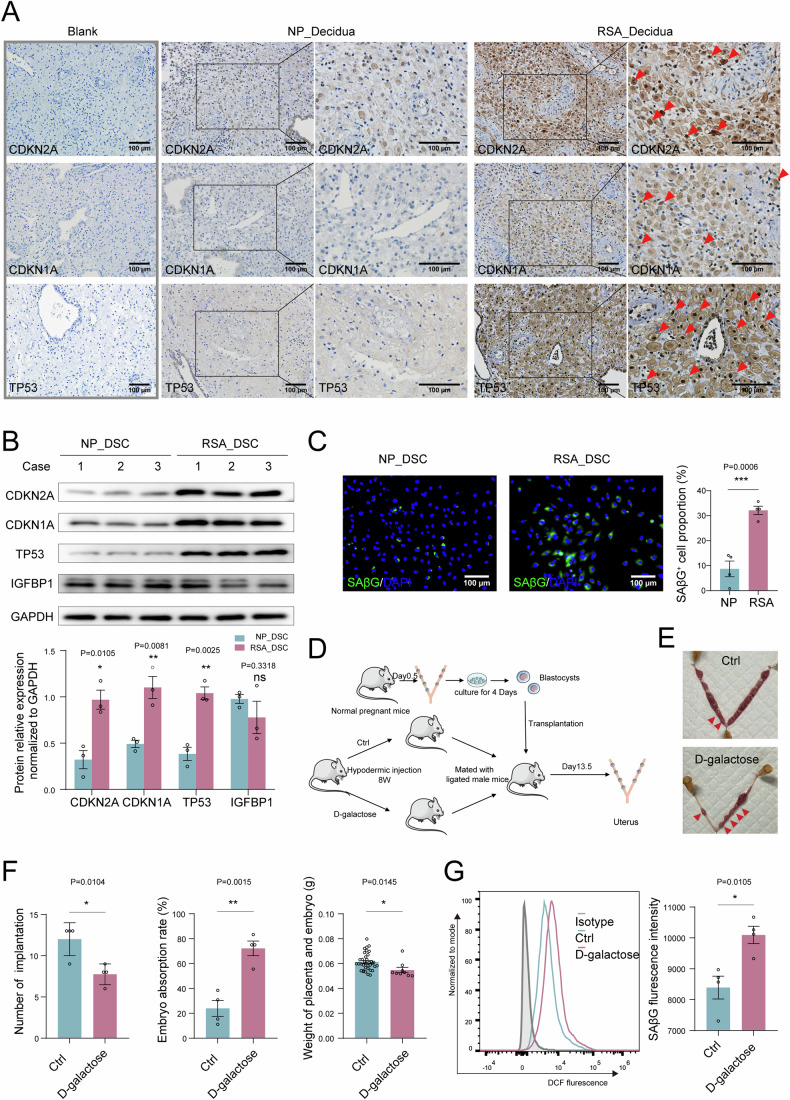
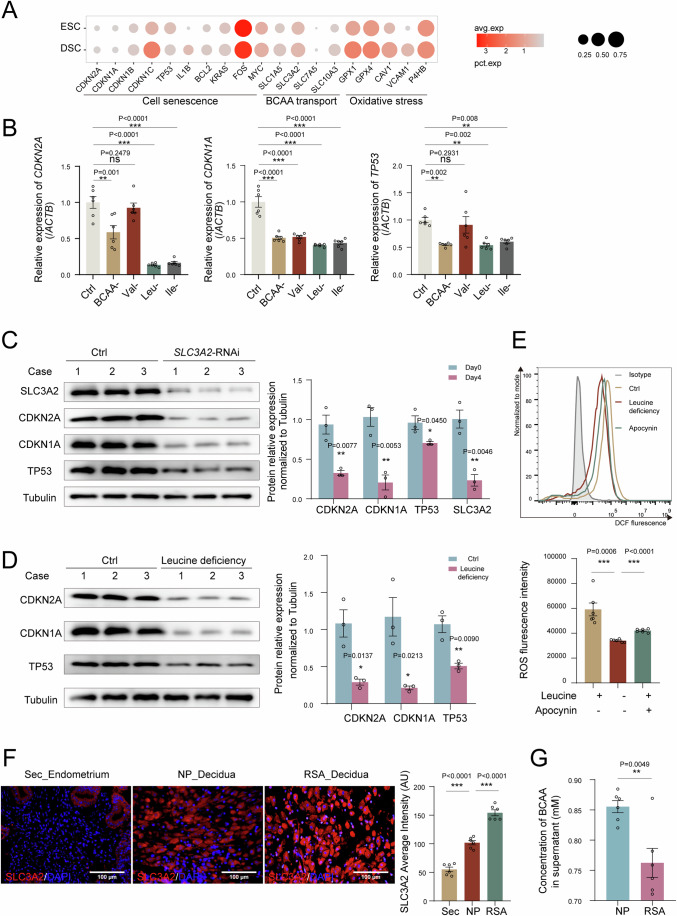
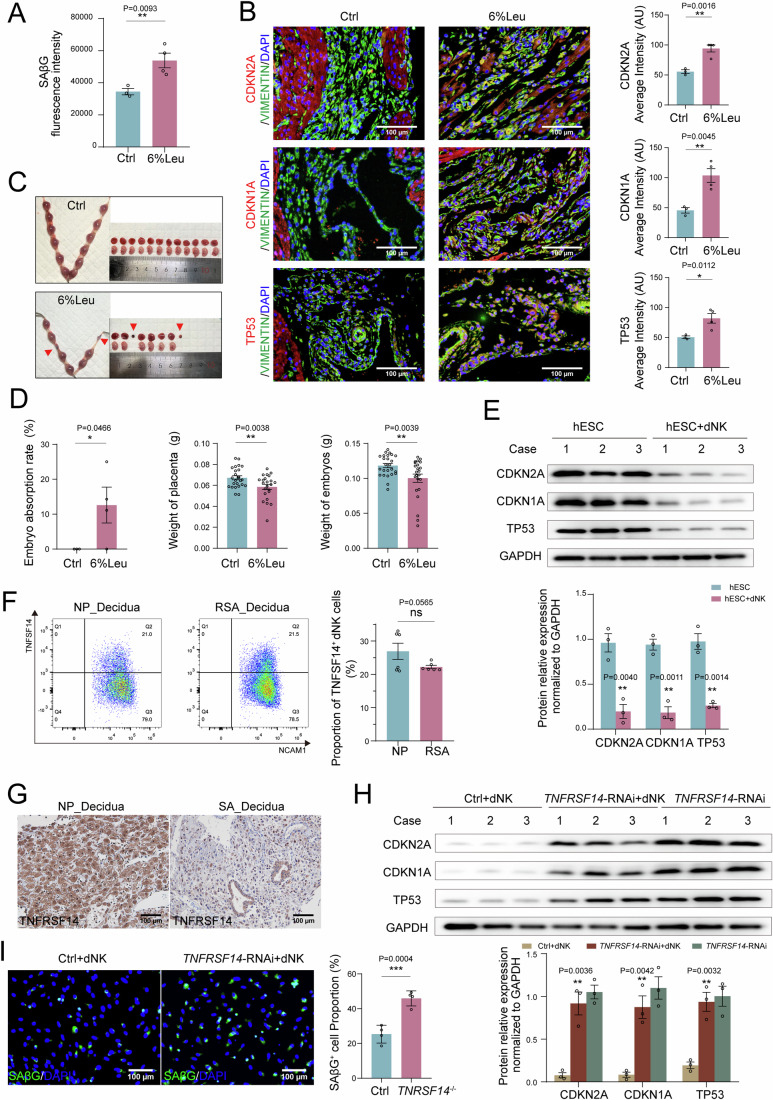
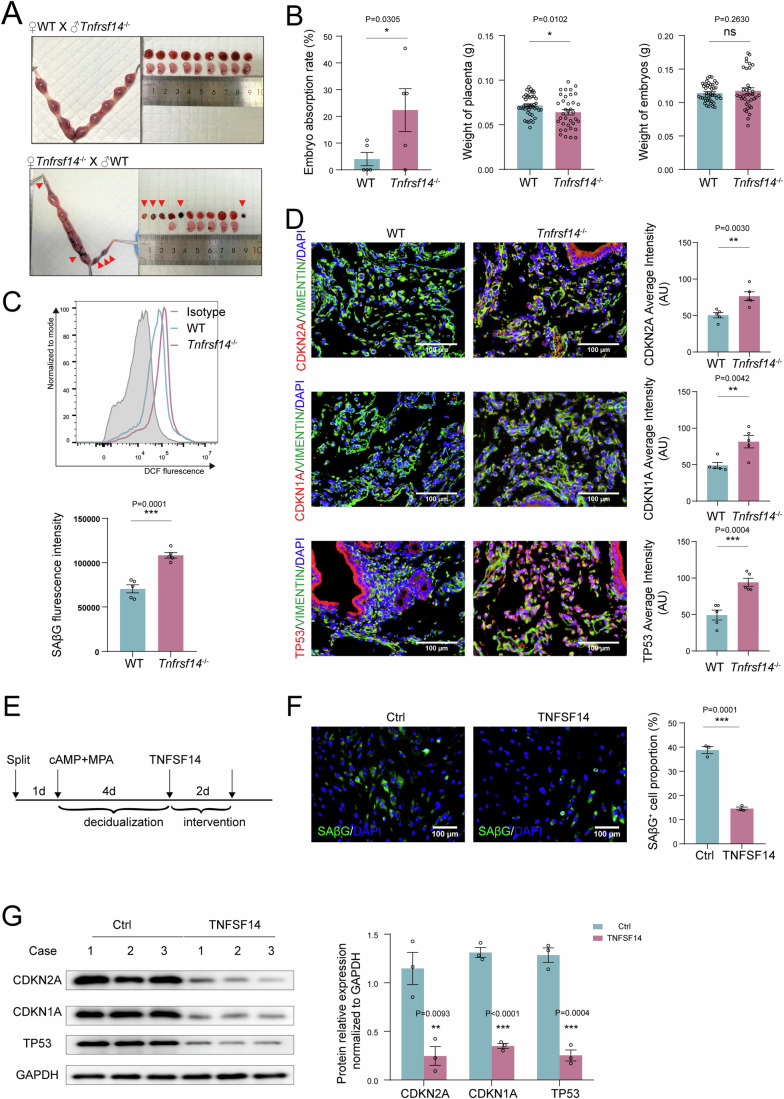
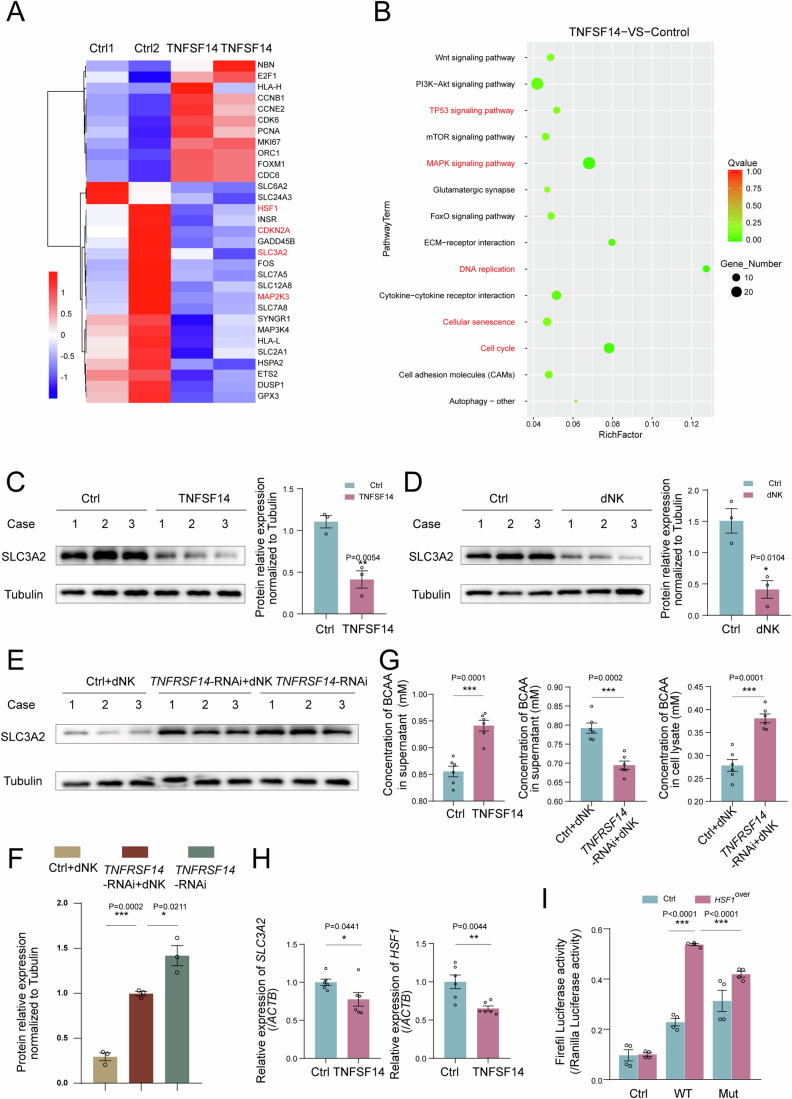
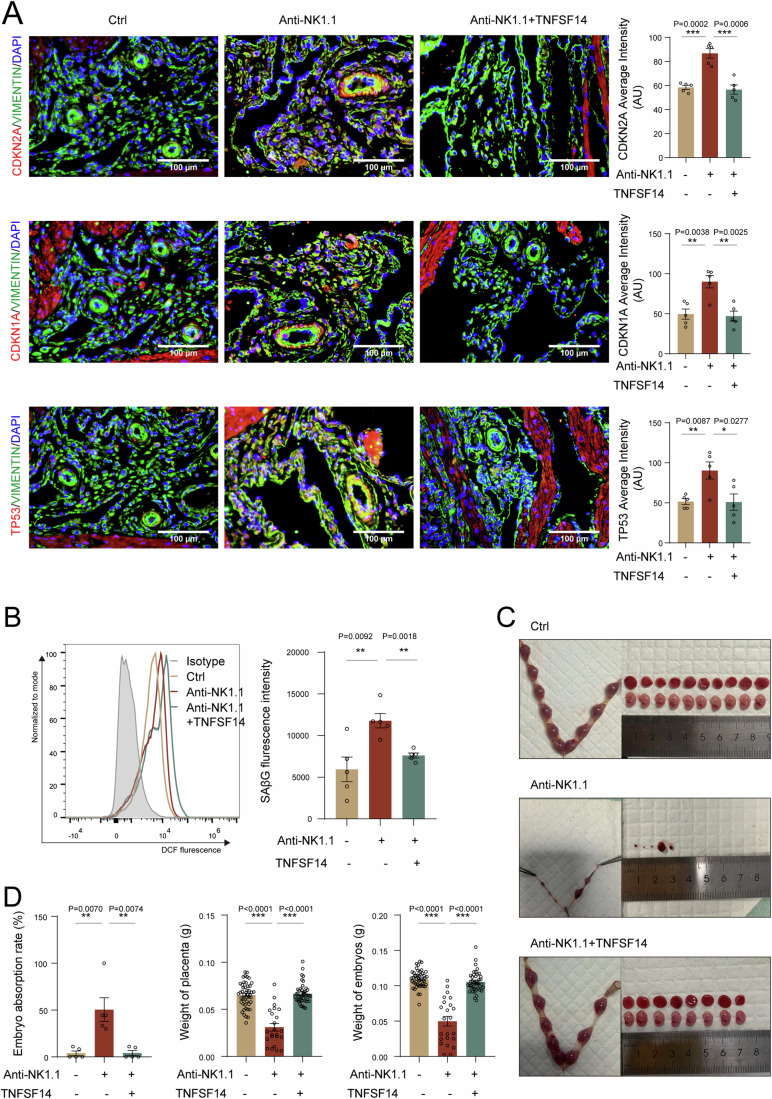
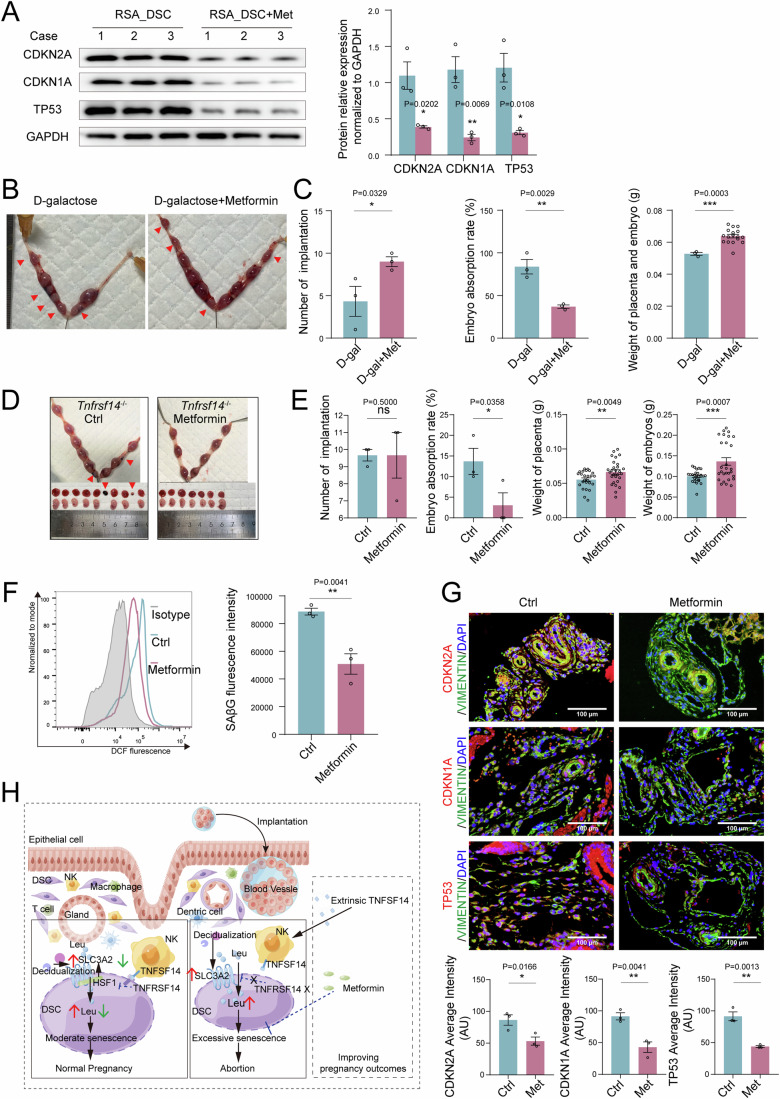
In preparation for a potential pregnancy, the endometrium of the uterus changes into a temporary structure called the decidua. Senescent decidual stromal cells (DSCs) are enriched in the decidua during decidualization, but the underlying mechanisms of this process remain unclear. Here, we performed single-cell RNA transcriptomics on ESCs and DSCs and found that cell senescence during decidualization is accompanied by increased levels of the branched-chain amino acid (BCAA) transporter SLC3A2. Depletion of leucine, one of the branched-chain amino acids, from cultured media decreased senescence, while high leucine diet resulted in increased senescence and high rates of embryo loss in mice. BCAAs induced senescence in DSCs via the p38 MAPK pathway. In contrast, TNFSF14+ decidual natural killer (dNK) cells were found to inhibit DSC senescence by interacting with its ligand TNFRSF14. As in mice fed high-leucine diets, both mice with NK cell depletion and Tnfrsf14-deficient mice with excessive uterine senescence experienced adverse pregnancy outcomes. Further, we found excessive uterine senescence, SLC3A2-mediated BCAA intake, and insufficient TNFRSF14 expression in the decidua of patients with recurrent spontaneous abortion. In summary, this study suggests that dNK cells maintain senescence homeostasis of DSCs via TNFSF14/TNFRSF14, providing a potential therapeutic strategy to prevent DSC senescence-associated spontaneous abortion.
Keywords: Abortion; Aging; Decidualization; Leucine; TNFRSF14.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- 32370914,92357306/MOST | National Natural Science Foundation of China (NSFC)
- 81901563/MOST | National Natural Science Foundation of China (NSFC)
- 82301895/MOST | National Natural Science Foundation of China (NSFC)
- 2023YFC2705403/MOST | National Key Research and Development Program of China (NKPs)
- 23ZR1408200/STCSM | Natural Science Foundation of Shanghai Municipality ()
LinkOut - more resources
Full Text Sources
Research Materials
