

A proteasome-dependent inhibition of SIRT-1 by the resveratrol analogue 4,4'-dihydroxy- trans- stilbene
- PMID: 39262665
- PMCID: PMC11384077
- DOI: 10.1016/j.jtcme.2024.03.001
A proteasome-dependent inhibition of SIRT-1 by the resveratrol analogue 4,4'-dihydroxy- trans- stilbene
Abstract
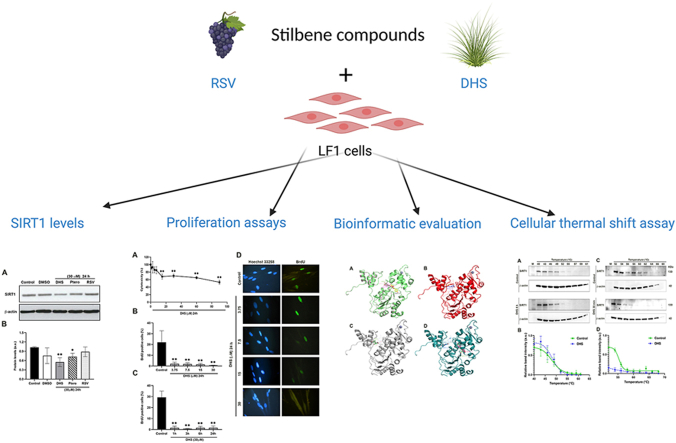
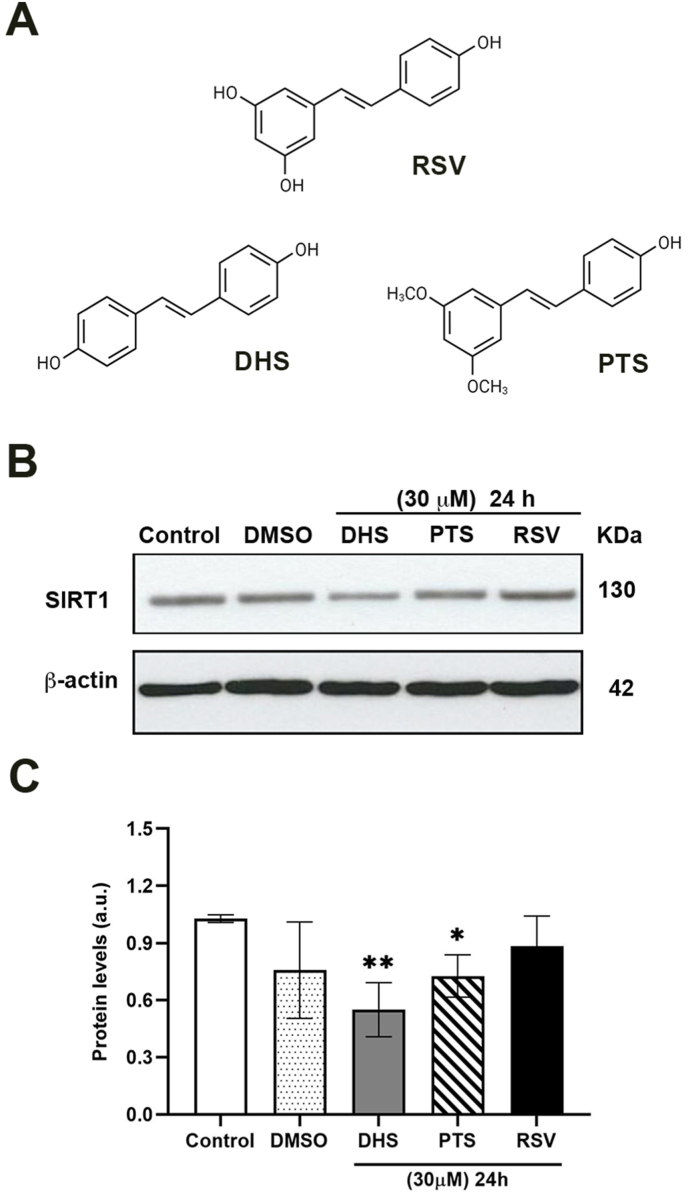
Background and aim: Resveratrol (RSV), is a stilbene-based compound exerting wide biological properties. Its analogue 4,4'-dihydroxy-trans-stilbene (DHS) has shown improved bioavailability and antiproliferative activity in vitro and in vivo. One of the hypotheses on how resveratrol works is based on SIRT1 activation. Since their strict structural similarities, we have explored a potential interaction between DHS and SIRT1, in comparison with the parental molecule.
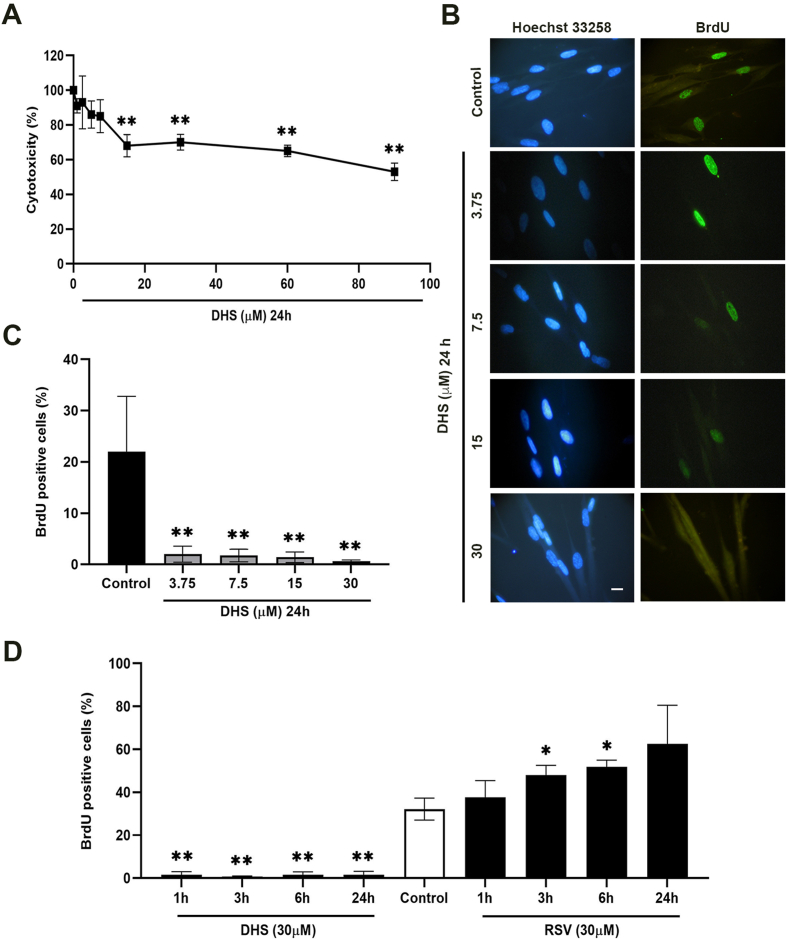
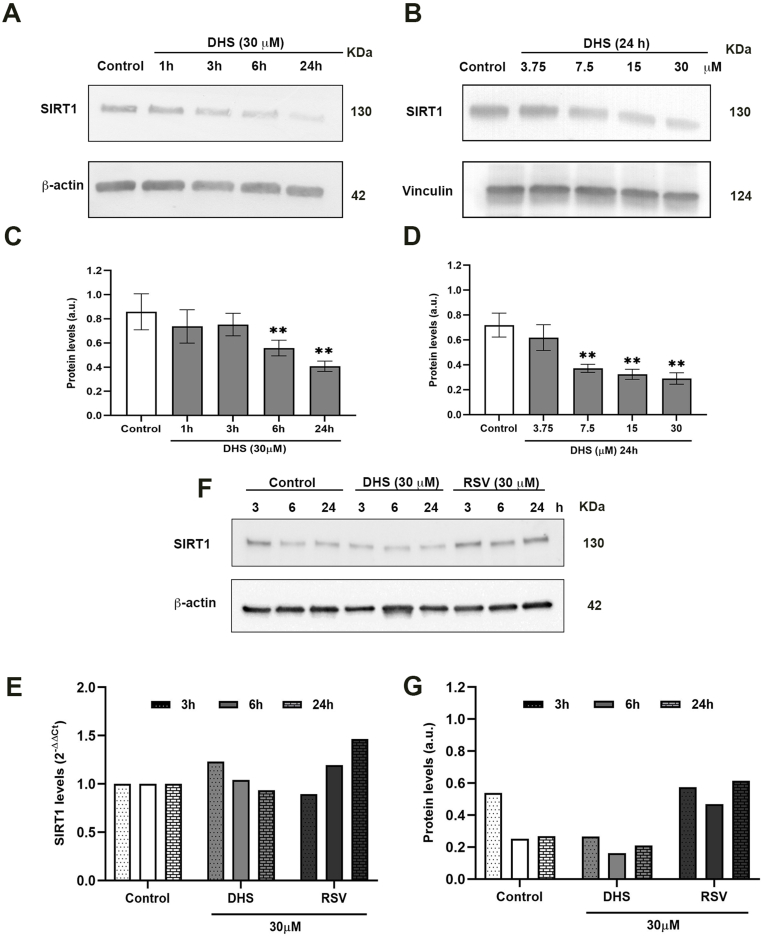
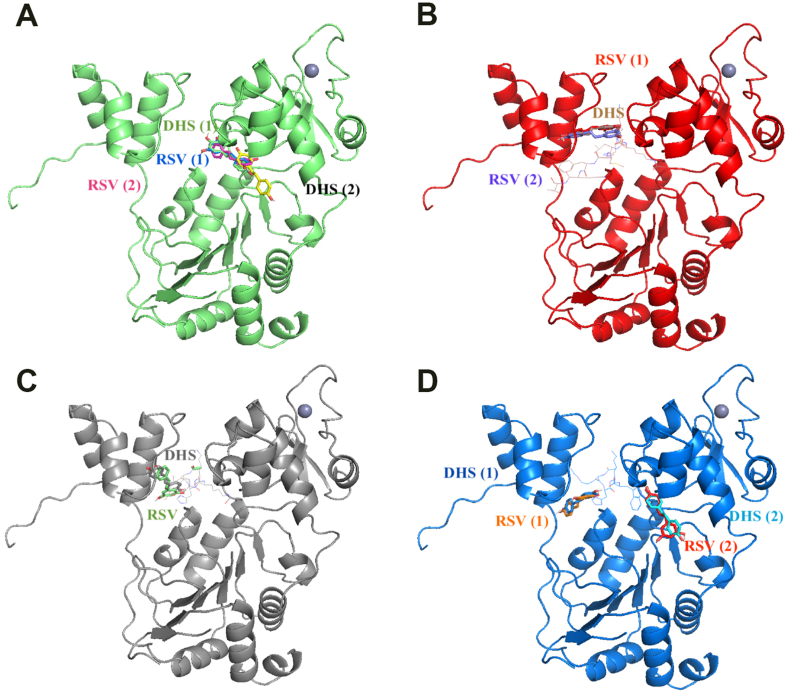
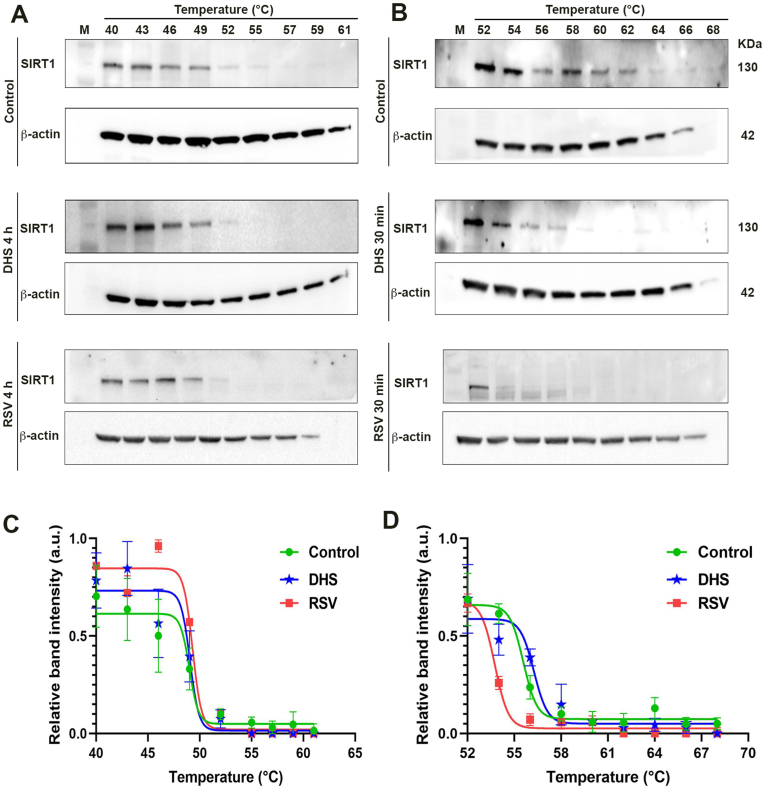
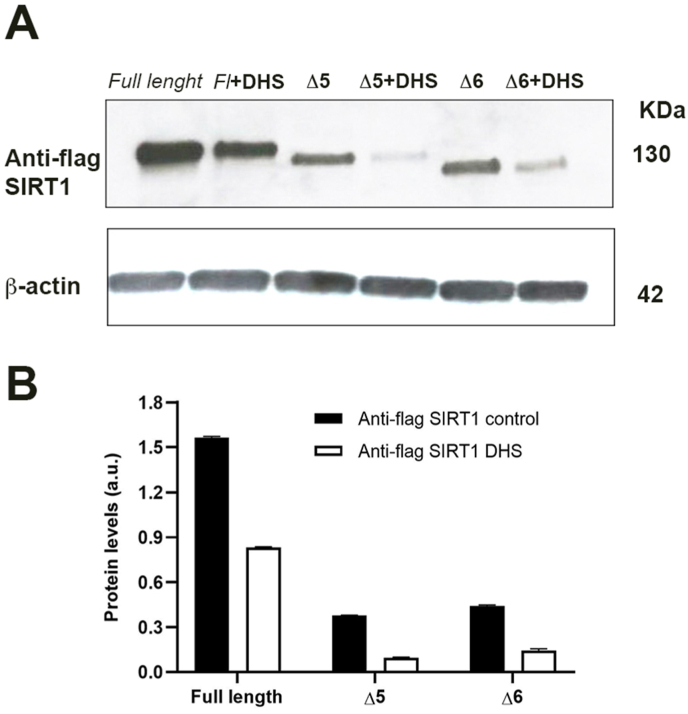
Experimental procedure: Timing of incubation and concentrations of DHS have been determined using MTT assay in normal human lung fibroblasts. Untreated, DHS- or RSV-treated cells were harvested and analysed by Western Blotting or RT-PCR, in order to evaluate SIRT1 levels/activity and expression, and by Cellular Thermal shift assay (CETSA) to check potential DHS or RSV-SIRT1 interaction. Transfection experiments have been performed with two SIRT1 mutants, based on the potential binding pockets identified by Molecular Docking analysis.
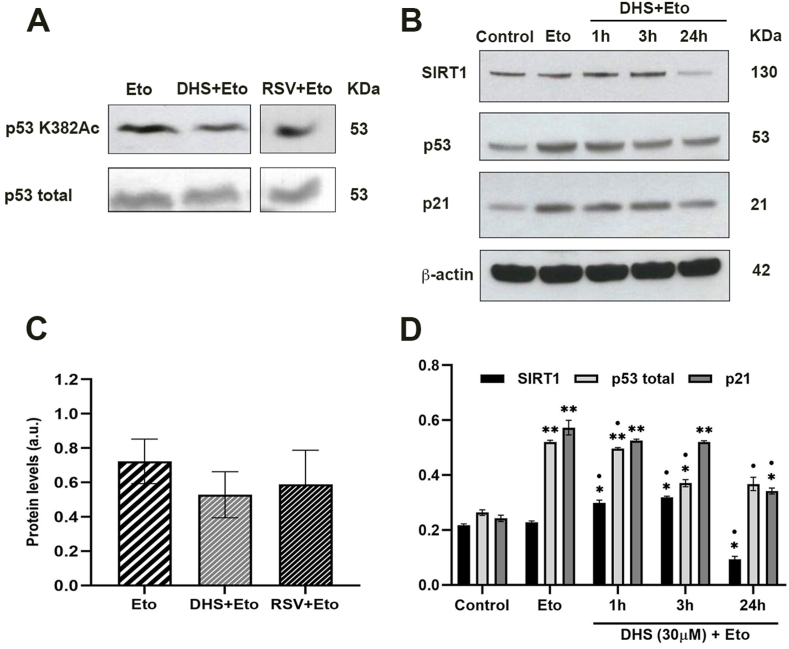
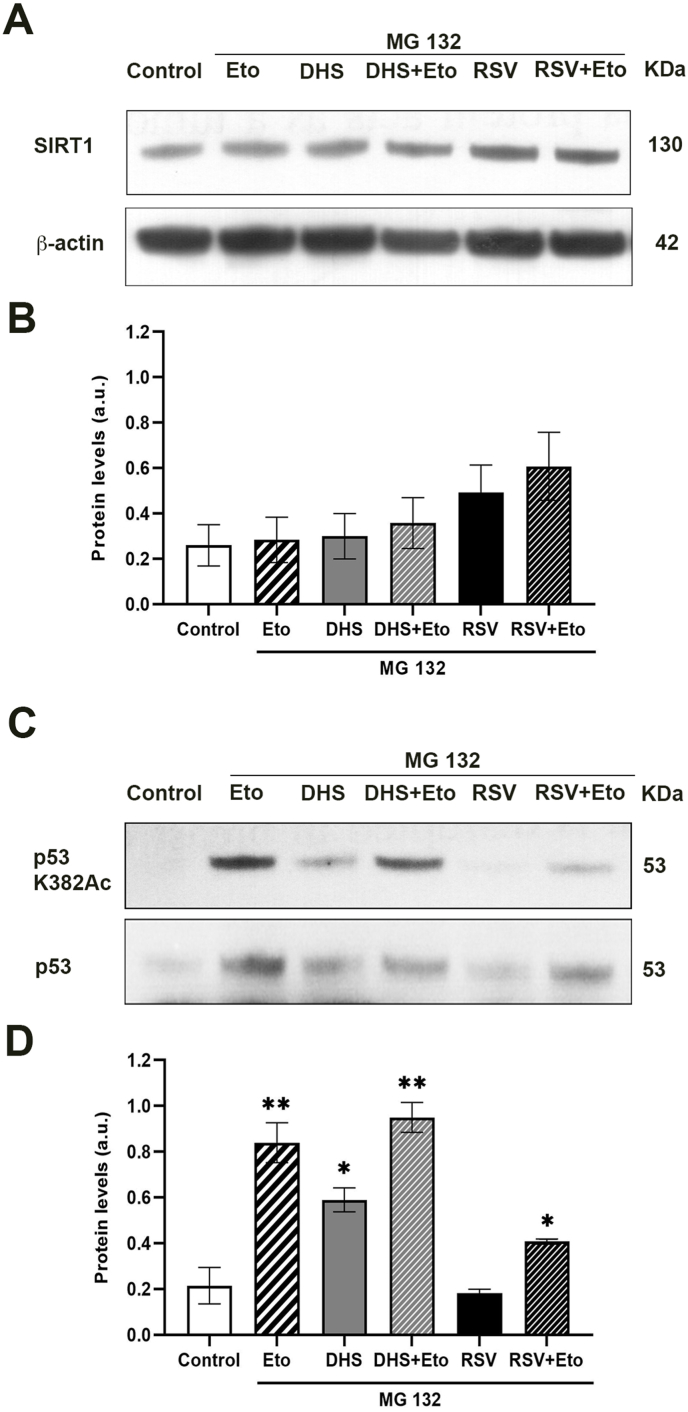
Results and conclusion: We unexpectedly found that DHS, but not RSV, exerted a time-dependent inhibitory effect on both SIRT1 protein levels and activity, the latter measured as p53 acetylation. At the mRNA level no significant changes were observed, whereas a proteasome-dependent mechanism was highlighted for the reduction of SIRT1 levels by DHS in experiments performed with the proteasome inhibitor MG132. Bioinformatics analysis suggested a higher affinity of RSV in binding all SIRT1 complexes compared to DHS, except comparable results for complex SIRT1-p53. Nevertheless, both CETSA and SIRT1 mutants transfected in cells did not confirm this interaction. In conclusion, DHS reduces SIRT1 protein level, thereby inhibiting its activity through a proteasome-mediated mechanism.
Keywords: 4,4′-dihydroxystilbene; Deacetylation; Resveratrol; Sirtuin-1.
© 2024 Center for Food and Biomolecules, National Taiwan University. Production and hosting by Elsevier Taiwan LLC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bradamante S., Barenghi L., Villa A. Cardiovascular protective effects of resveratrol. Cardiovasc Drug Rev. 2004;22:169–188. - PubMed
-
- Soleas G.J., Diamandis E.P., Goldberg D.M. The world of resveratrol. Adv Exp Med Biol. 2001;492:159–182. - PubMed
-
- Hazafa A., Iqbal M.O., Javaid U., et al. Inhibitory effect of polyphenols (phenolic acids, lignans, and stilbenes) on cancer by regulating signal transduction pathways: a review. Clin Transl Oncol. 2022;24:432–445. - PubMed
-
- Kang J.H., Park Y.H., Choi S.W., Yang E.K., Lee W.J. Resveratrol derivatives potently induce apoptosis in human promyelocytic leukemia cells. Exp Mol Med. 2003;35:467–474. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous