

Maternal high-fat diet alters Tet-mediated epigenetic regulation during heart development
- PMID: 39262804
- PMCID: PMC11388159
- DOI: 10.1016/j.isci.2024.110631
Maternal high-fat diet alters Tet-mediated epigenetic regulation during heart development
Abstract
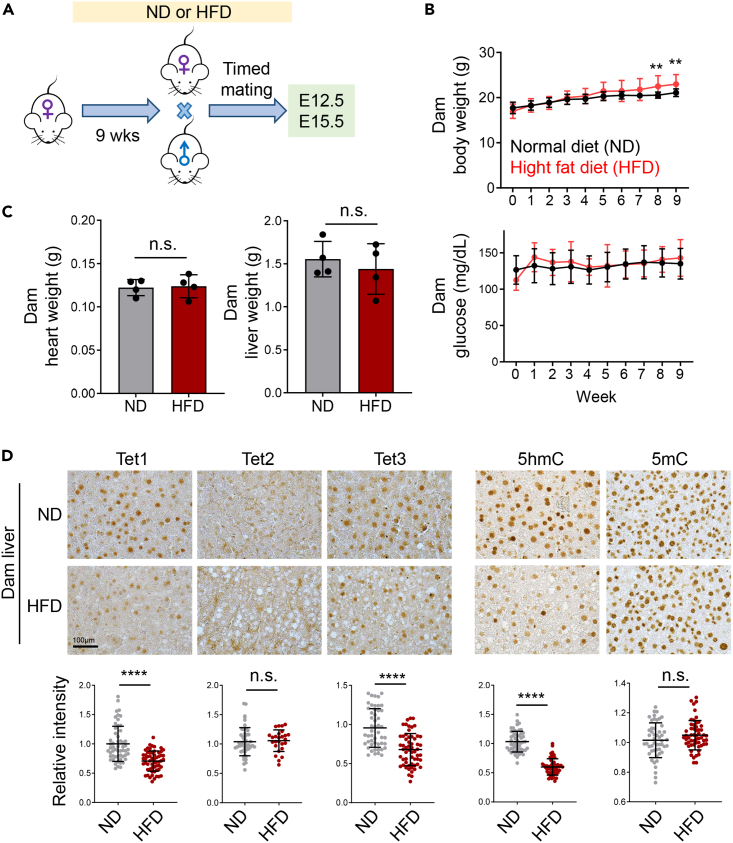
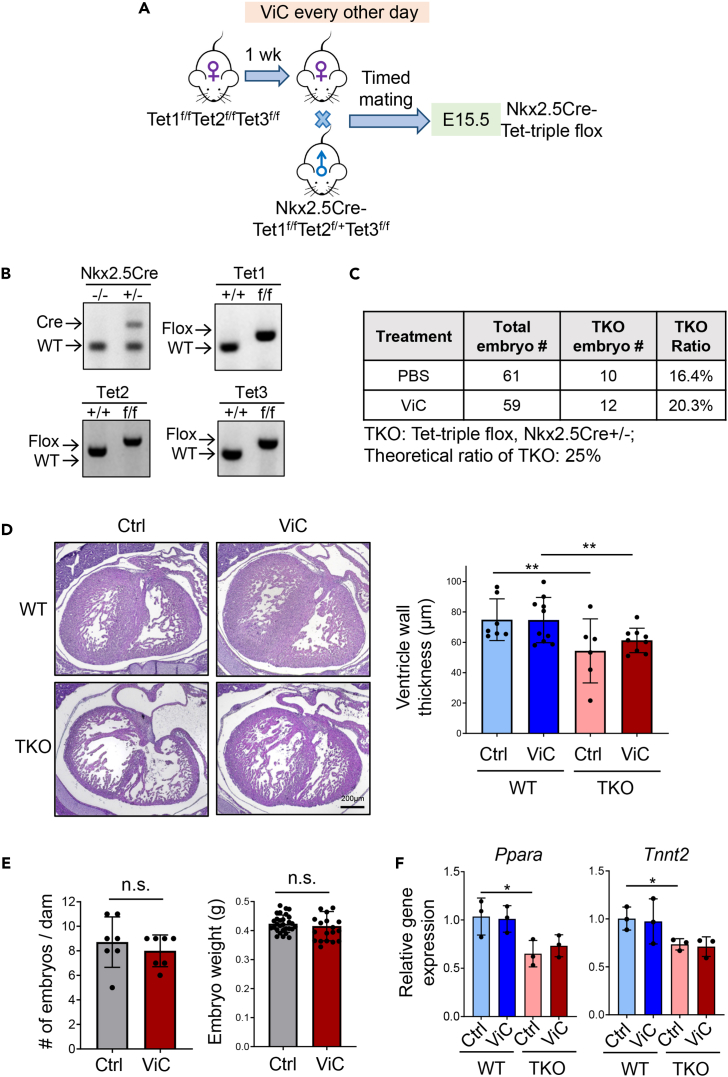
Imbalanced dietary intake, such as a high-fat diet (HFD) during pregnancy, has been associated with adverse offspring outcomes. Metabolic stress from imbalanced food intake alters the function of epigenetic regulators, resulting in abnormal transcriptional outputs in embryos to cause congenital disorders. We report herein that maternal HFD exposure causes metabolic changes in pregnant mice and non-compaction cardiomyopathy (NCC) in E15.5 embryos, accompanied by decreased 5-hydroxymethylcytosine (5hmC) levels and altered chromatin accessibility in embryonic heart tissues. Remarkably, maternal vitamin C supplementation mitigates these detrimental effects, likely by restoring iron, a cofactor for Tet enzymes, in a reduced state. Using a genetic approach, we further demonstrated that the cardioprotective benefits of vitamin C under HFD conditions are attributable to enhanced Tet activity. Our results highlight an interaction between maternal diet, specifically HFD or vitamin C, and epigenetic modifications during early heart development, emphasizing the importance of balanced maternal nutrition for healthy embryonic development.
Keywords: Developmental biology; Diet; Epigenetics; Model organism; Pregnancy.
© 2024 The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Mdaki K.S., Larsen T.D., Wachal A.L., Schimelpfenig M.D., Weaver L.J., Dooyema S.D.R., Louwagie E.J., Baack M.L. Maternal high-fat diet impairs cardiac function in offspring of diabetic pregnancy through metabolic stress and mitochondrial dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2016;310:H681–H692. doi: 10.1152/ajpheart.00795.2015. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
