

Brain shapes of large-bodied, flightless ratites (Aves: Palaeognathae) emerge through distinct developmental allometries
- PMID: 39263457
- PMCID: PMC11387061
- DOI: 10.1098/rsos.240765
Brain shapes of large-bodied, flightless ratites (Aves: Palaeognathae) emerge through distinct developmental allometries
Abstract
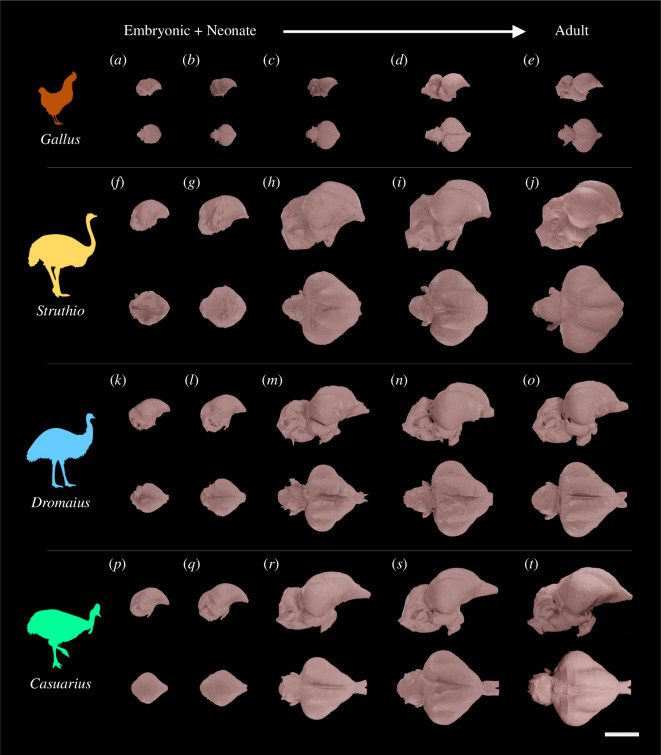
Comparative neuroanatomical studies have long debated the role of development in the evolution of novel and disparate brain morphologies. Historically, these studies have emphasized whether evolutionary shifts along conserved or distinct developmental allometric trends cause changes in brain morphologies. However, the degree to which interspecific differences between variably sized taxa originate through modifying developmental allometry remains largely untested. Taxa with disparate brain shapes and sizes thus allow for investigation into how developmental trends contribute to neuroanatomical diversification. Here, we examine a developmental series of large-bodied ratite birds (approx. 60-140 kg). We use three-dimensional geometric morphometrics on cephalic endocasts of common ostriches, emus and southern cassowaries and compare their developmental trajectories with those of the more modestly sized domestic chicken, previously shown to be in the same allometric grade as ratites. The results suggest that ratites and chickens exhibit disparate endocranial shapes not simply accounted for by their size differences. When shape and age are examined, chickens partly exhibit more accelerated and mature brain shapes than ratites of similar size and age. Taken together, our study indicates that disparate brain shapes between these differently sized taxa have emerged from the evolution of distinct developmental allometries, rather than simply following conserved scaling trends.
Keywords: Palaeognathae; allometry; endocasts; geometric morphometrics; micro-CT imaging; ontogeny.
© 2024 The Author(s).
Conflict of interest statement
We declare we have no competing interests.
Figures





Similar articles
-
Distinct developmental pathways underlie independent losses of flight in ratites.Biol Lett. 2017 Jul;13(7):20170234. doi: 10.1098/rsbl.2017.0234. Biol Lett. 2017. PMID: 28679696 Free PMC article.
-
Ratite nonmonophyly: independent evidence from 40 novel Loci.Syst Biol. 2013 Jan 1;62(1):35-49. doi: 10.1093/sysbio/sys067. Epub 2012 Jul 24. Syst Biol. 2013. PMID: 22831877
-
Novel neuroanatomical integration and scaling define avian brain shape evolution and development.Elife. 2021 Jul 6;10:e68809. doi: 10.7554/eLife.68809. Elife. 2021. PMID: 34227464 Free PMC article.
-
Neoteny and the thyroid in ratites.Rev Reprod. 1996 May;1(2):78-81. doi: 10.1530/ror.0.0010078. Rev Reprod. 1996. PMID: 9414443 Review.
-
An updated look at the mating system, parental care and androgen seasonal variations in ratites.Gen Comp Endocrinol. 2022 Jul 1;323-324:114034. doi: 10.1016/j.ygcen.2022.114034. Epub 2022 Mar 31. Gen Comp Endocrinol. 2022. PMID: 35367461 Review.
Cited by
-
Dinosaur palaeoneurology: an evolving science.Biol Lett. 2024 Dec;20(12):20240472. doi: 10.1098/rsbl.2024.0472. Epub 2024 Dec 18. Biol Lett. 2024. PMID: 39689851 Free PMC article. Review.
References
-
- Striedter GF. 2004. Brain evolution. In The human nervous system (eds Paxinos G, Mai JK), pp. 3–21, 2nd edn. London, UK: Elsevier. (10.1016/B978-012547626-3/50002-8) - DOI
-
- Rilling JK. 2006. Human and nonhuman primate brains: are they allometrically scaled versions of the same design? Evol. Anthropol. 15, 65–77. (10.1002/evan.20095) - DOI
Associated data
LinkOut - more resources
Full Text Sources
