

Phenotypic profiling of human induced regulatory T cells at early differentiation: insights into distinct immunosuppressive potential
- PMID: 39264416
- PMCID: PMC11393232
- DOI: 10.1007/s00018-024-05429-3
Phenotypic profiling of human induced regulatory T cells at early differentiation: insights into distinct immunosuppressive potential
Abstract
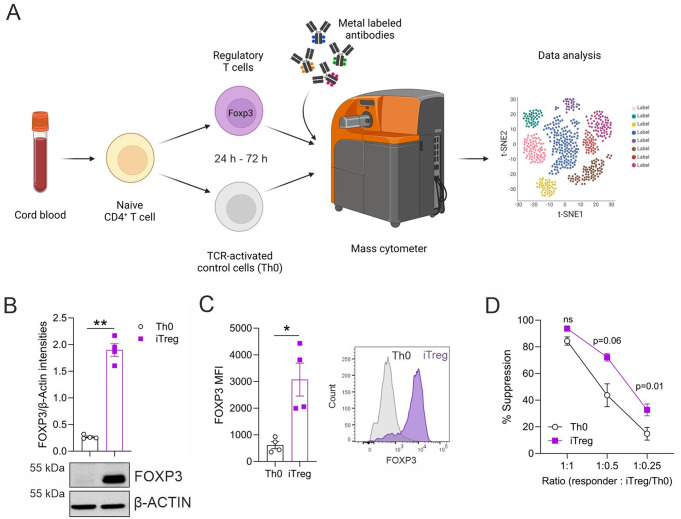
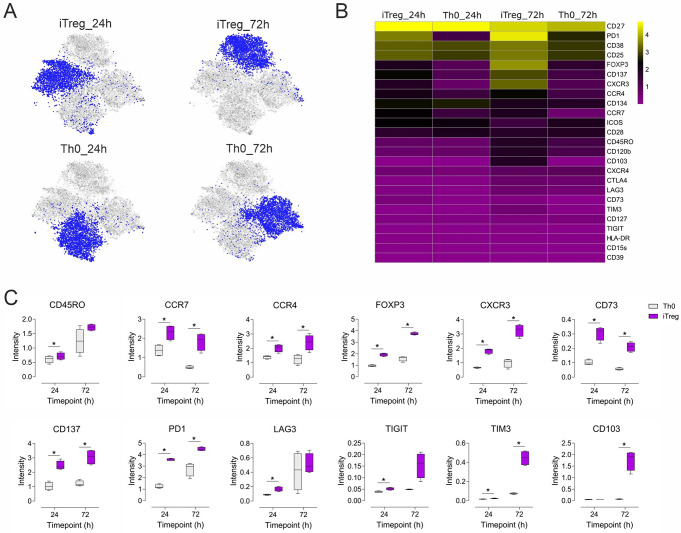
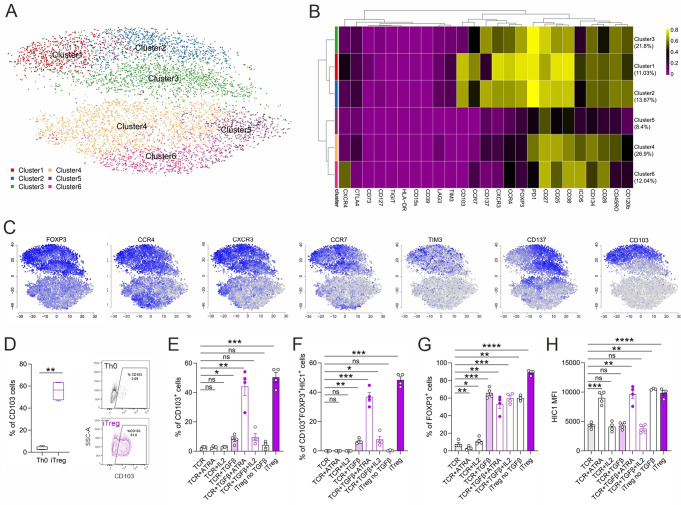
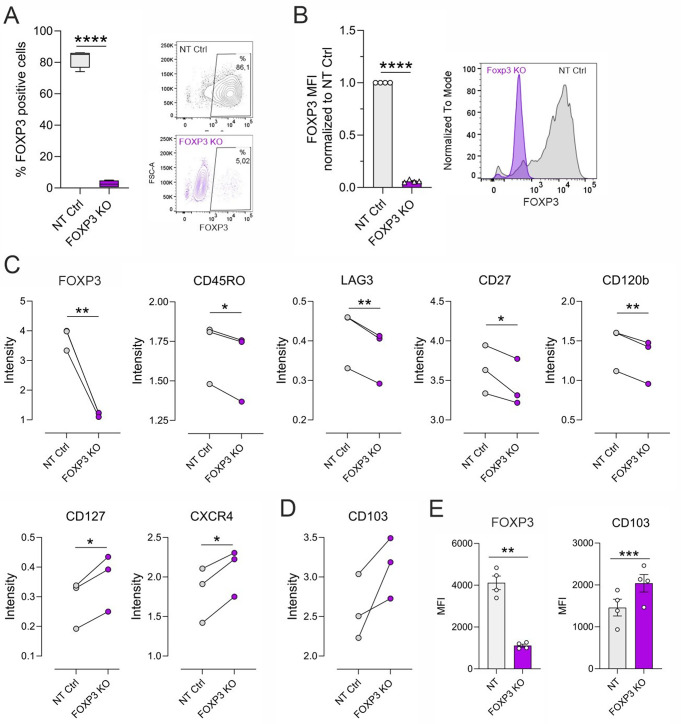
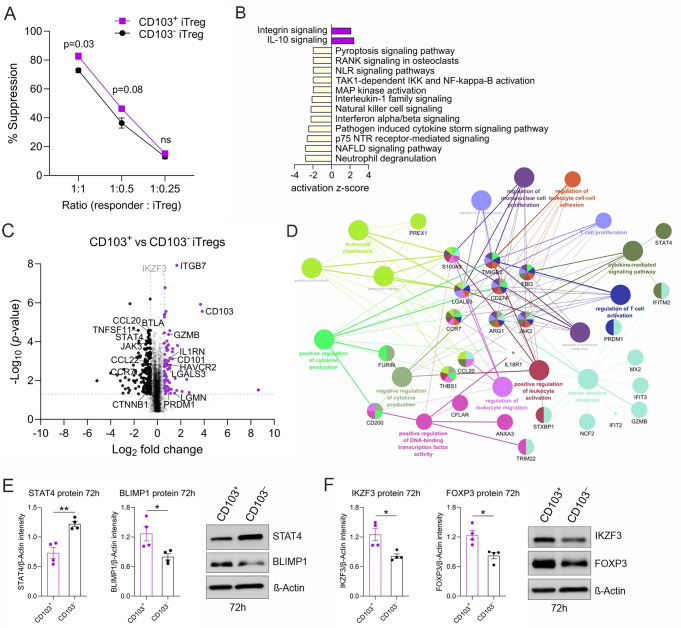
Regulatory T cells (Tregs) play a key role in suppressing systemic effector immune responses, thereby preventing autoimmune diseases but also potentially contributing to tumor progression. Thus, there is great interest in clinically manipulating Tregs, but the precise mechanisms governing in vitro-induced Treg (iTreg) differentiation are not yet fully understood. Here, we used multiparametric mass cytometry to phenotypically profile human iTregs during the early stages of in vitro differentiation at single-cell level. A panel of 25 metal-conjugated antibodies specific to markers associated with human Tregs was used to characterize these immunomodulatory cells. We found that iTregs highly express the transcription factor FOXP3, as well as characteristic Treg-associated surface markers (e.g. CD25, PD1, CD137, CCR4, CCR7, CXCR3, and CD103). Expression of co-inhibitory factors (e.g. TIM3, LAG3, and TIGIT) increased slightly at late stages of iTreg differentiation. Further, CD103 was upregulated on a subpopulation of iTregs with greater suppressive capacity than their CD103- counterparts. Using mass-spectrometry-based proteomics, we showed that sorted CD103+ iTregs express factors associated with immunosuppression. Overall, our study highlights that during early stages of differentiation, iTregs resemble memory-like Treg features with immunosuppressive activity, and provides opportunities for further investigation into the molecular mechanisms underlying Treg function.
Keywords: CD103; Differentiation; FOXP3; Mass cytometry; Mass spectrometry; Regulatory T cells.
© 2024. The Author(s).
Conflict of interest statement
A.M. is a cofounder of Site Tx, Arsenal Biosciences, Spotlight Therapeutics and Survey Genomics, serves on the boards of directors at Site Tx, Spotlight Therapeutics and Survey Genomics, is a member of the scientific advisory boards of Site Tx, Arsenal Biosciences, Spotlight Therapeutics, Survey Genomics, NewLimit, Amgen, and Tenaya, owns stock in Arsenal Biosciences, Site Tx, Spotlight Therapeutics, NewLimit, Survey Genomics, Tenaya and Lightcast and has received fees from Site Tx, Arsenal Biosciences, Spotlight Therapeutics, NewLimit, 23andMe, PACT Pharma, Juno Therapeutics, Tenaya, Lightcast, Trizell, Vertex, Merck, Amgen, Genentech, GLG, ClearView Healthcare, AlphaSights, Rupert Case Management, Bernstein and ALDA. A.M. is an investor in and informal advisor to Offline Ventures and a client of EPIQ. The Marson laboratory has received research support from the Parker Institute for Cancer Immunotherapy, the Emerson Collective, Juno Therapeutics, Epinomics, Sanofi, GlaxoSmithKline, Gilead and Anthem and reagents from Genscript and Illumina.The other authors declare no competing interests.
Figures
References
-
- Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M (1995) Immunologic self-tolerance maintained by activated T cells expressing IL-2 receptor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol 155 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
