

Unraveling the phenotypic states of human innate-like T cells: Comparative insights with conventional T cells and mouse models
- PMID: 39264810
- PMCID: PMC11552652
- DOI: 10.1016/j.celrep.2024.114705
Unraveling the phenotypic states of human innate-like T cells: Comparative insights with conventional T cells and mouse models
Abstract
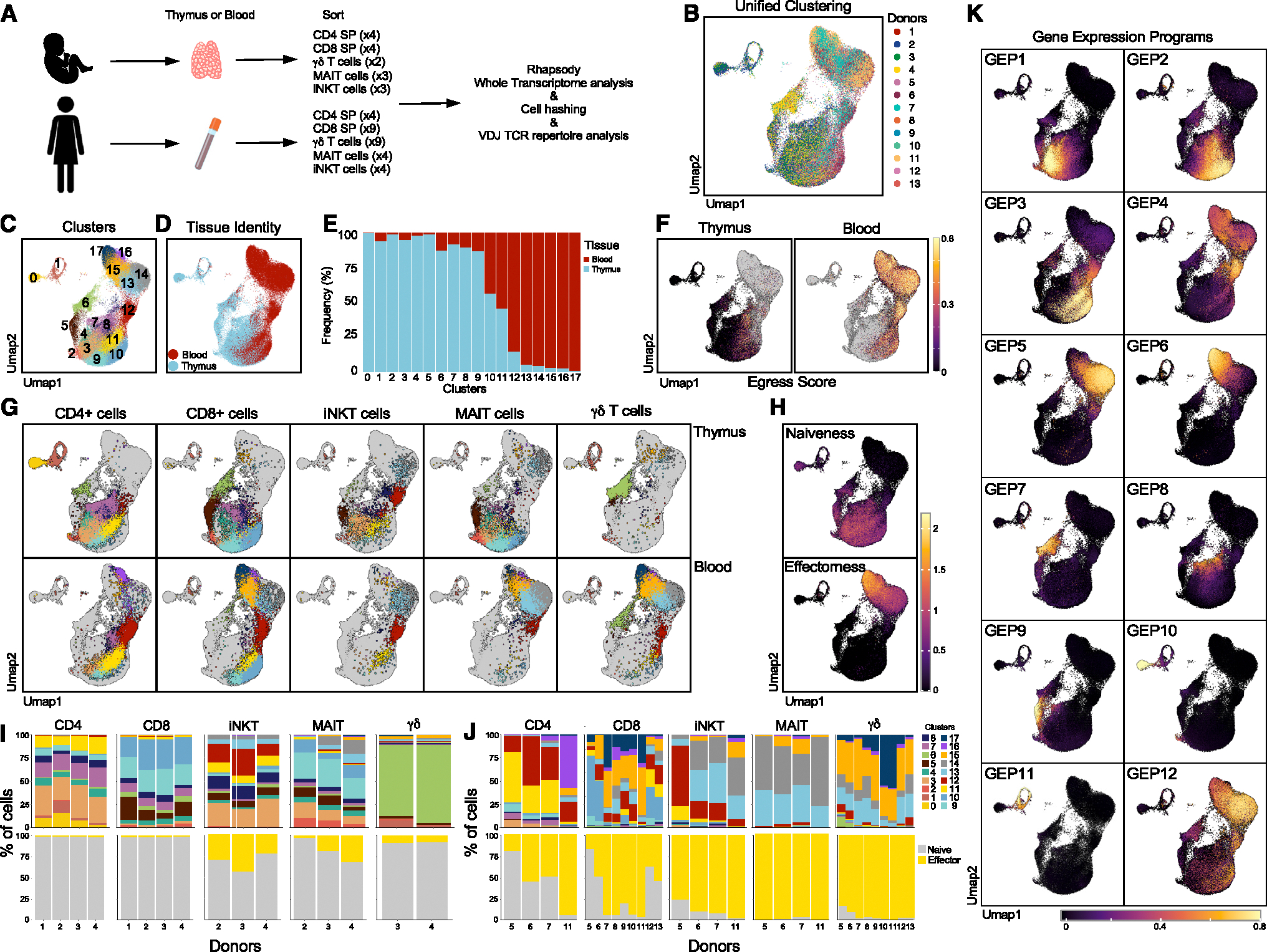
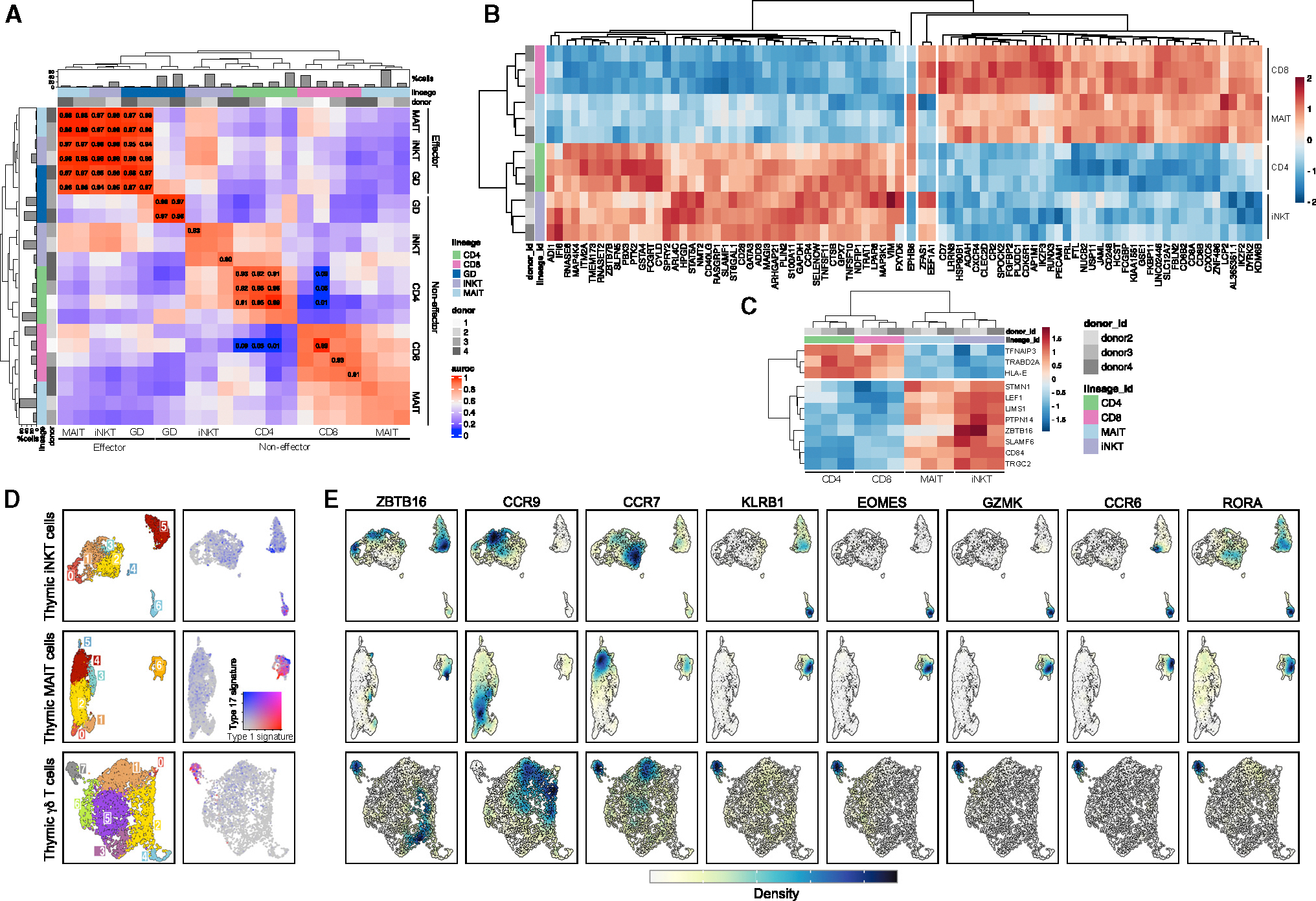
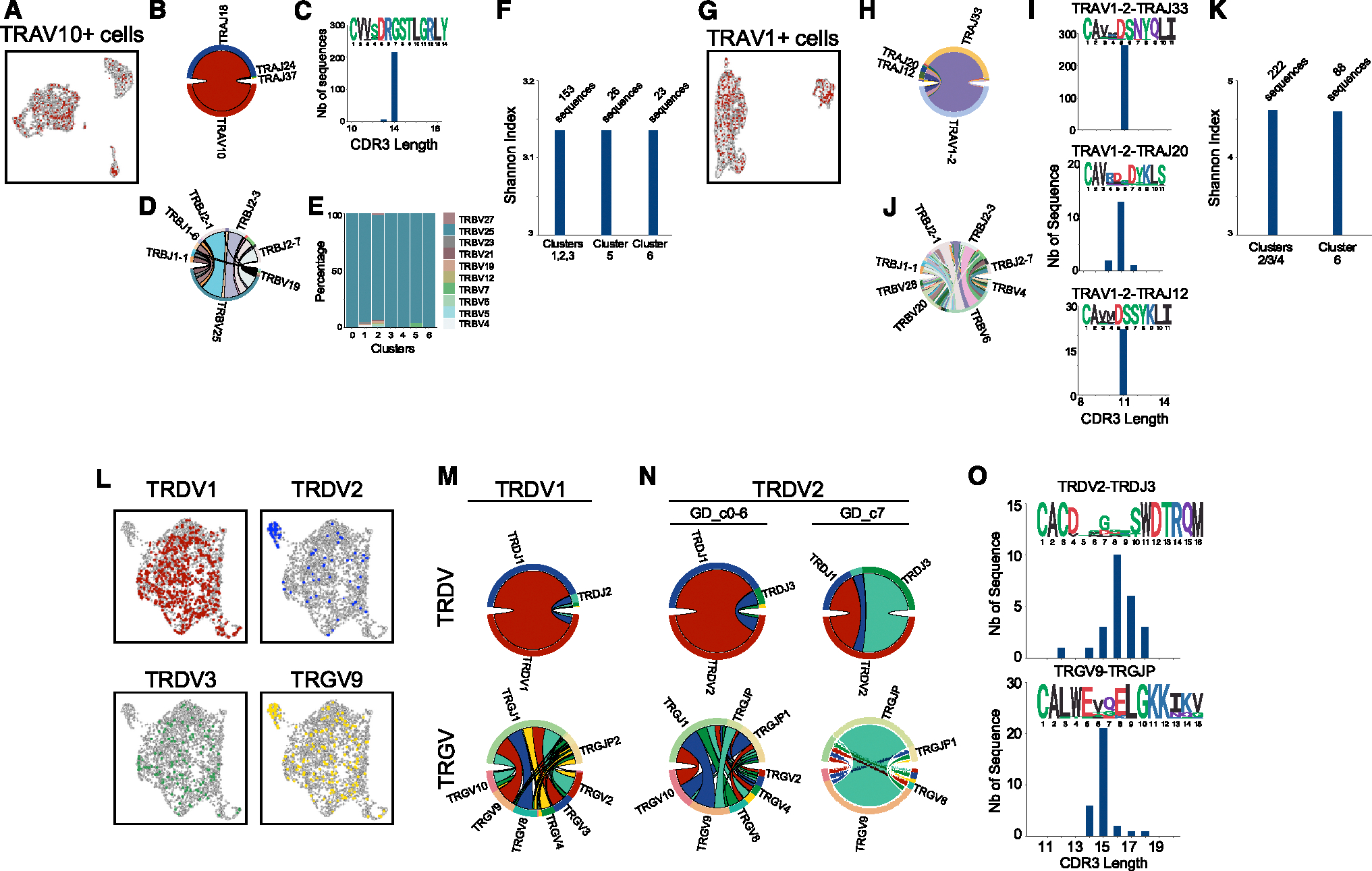
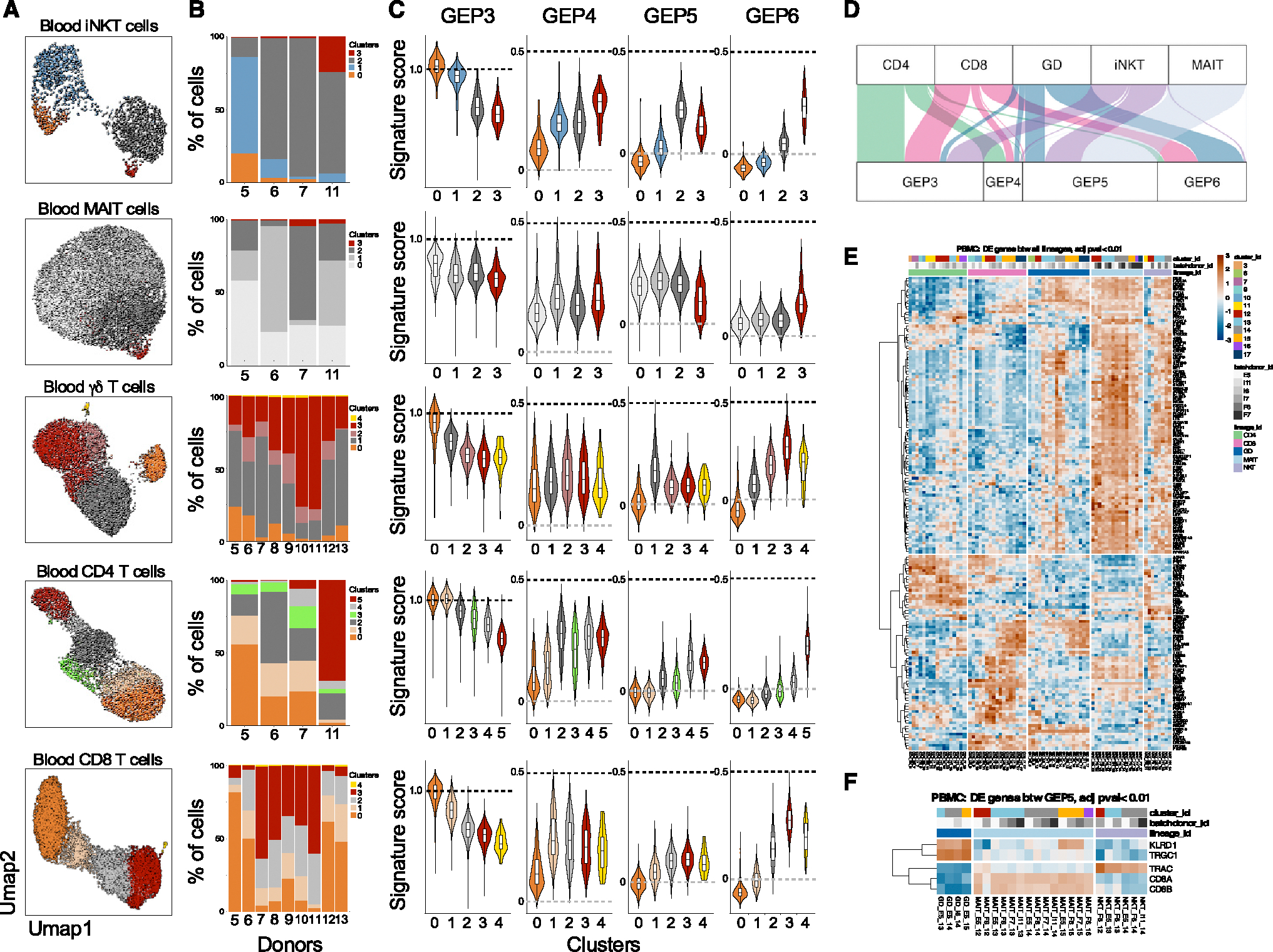
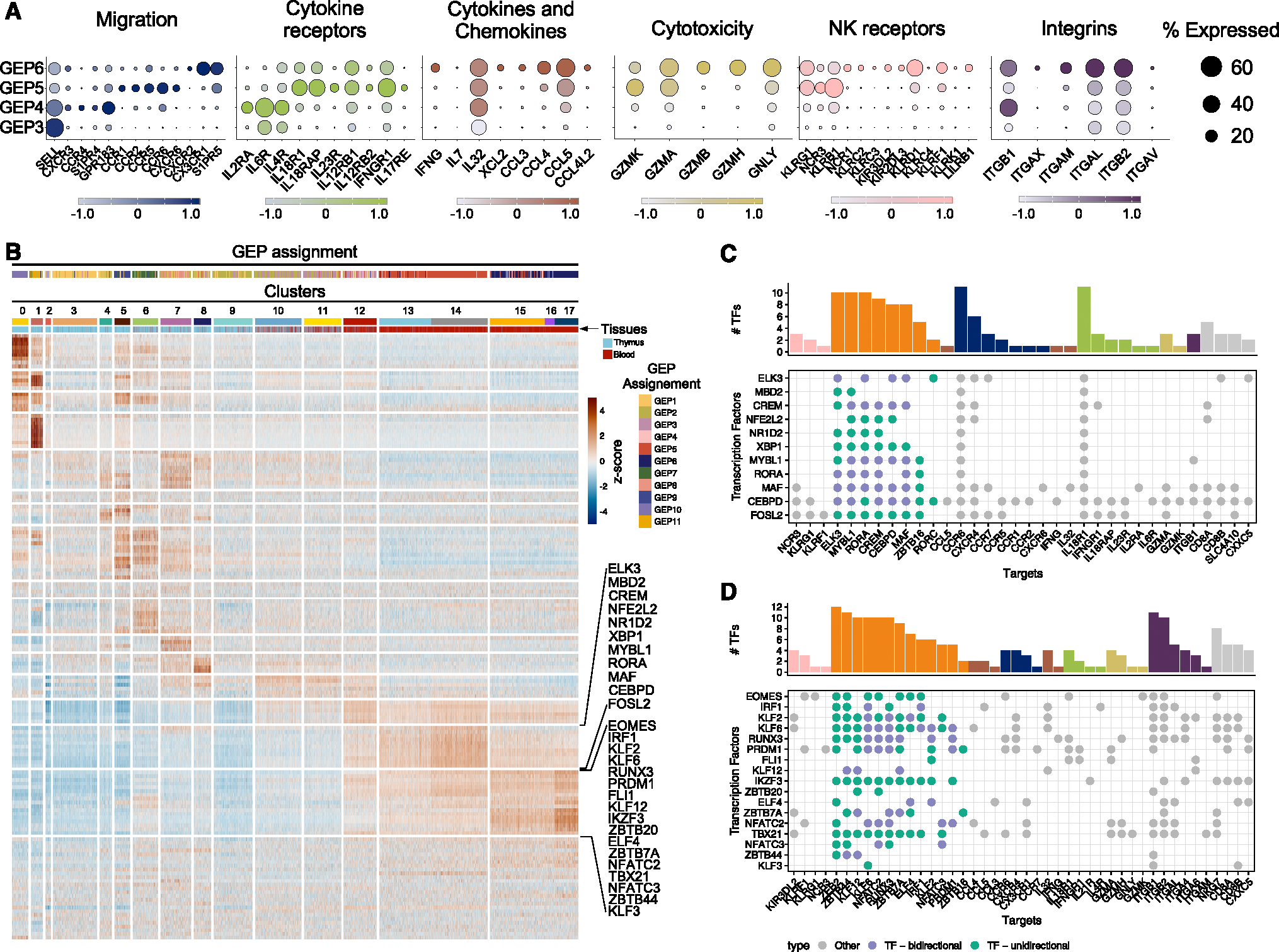
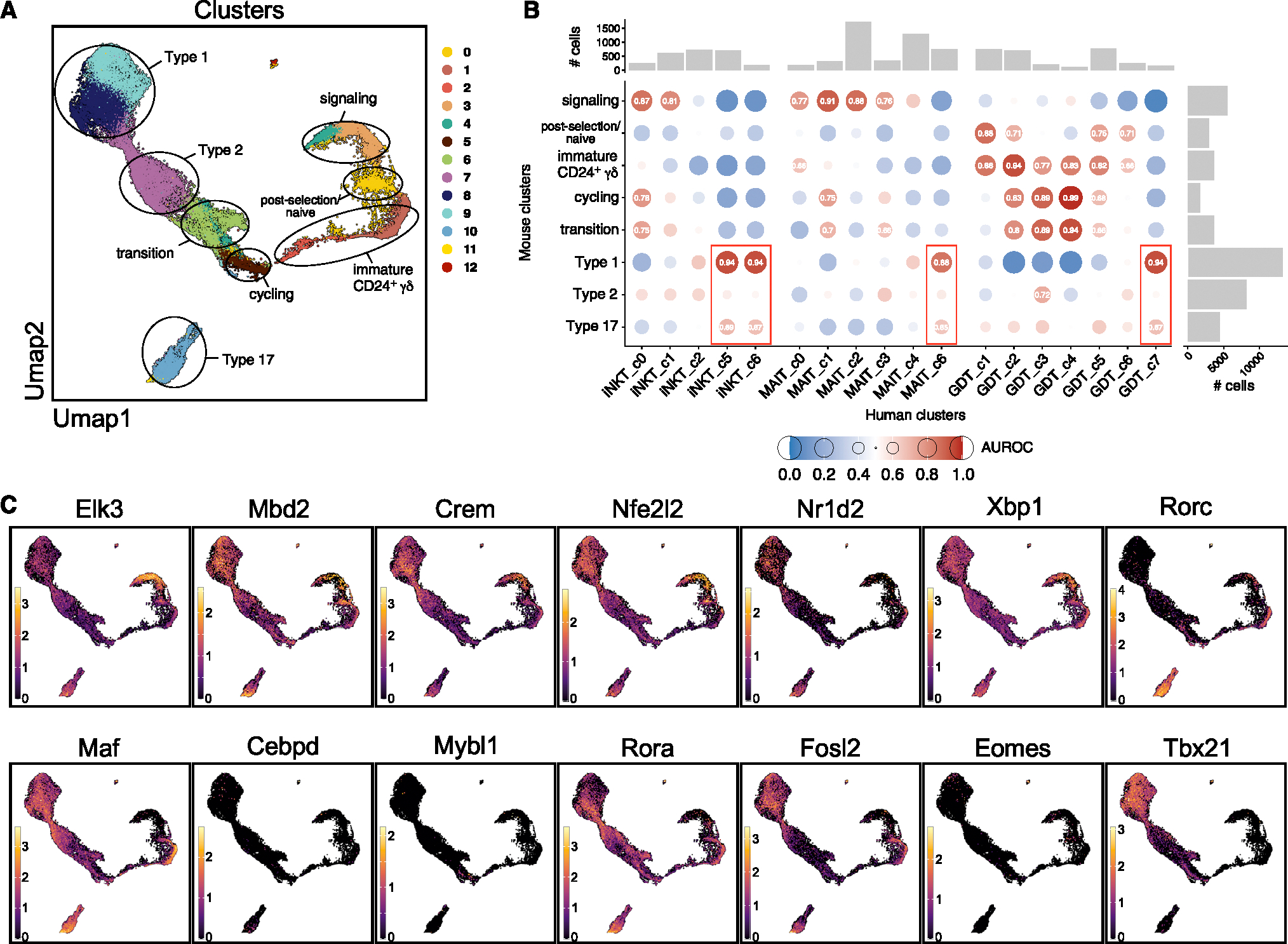
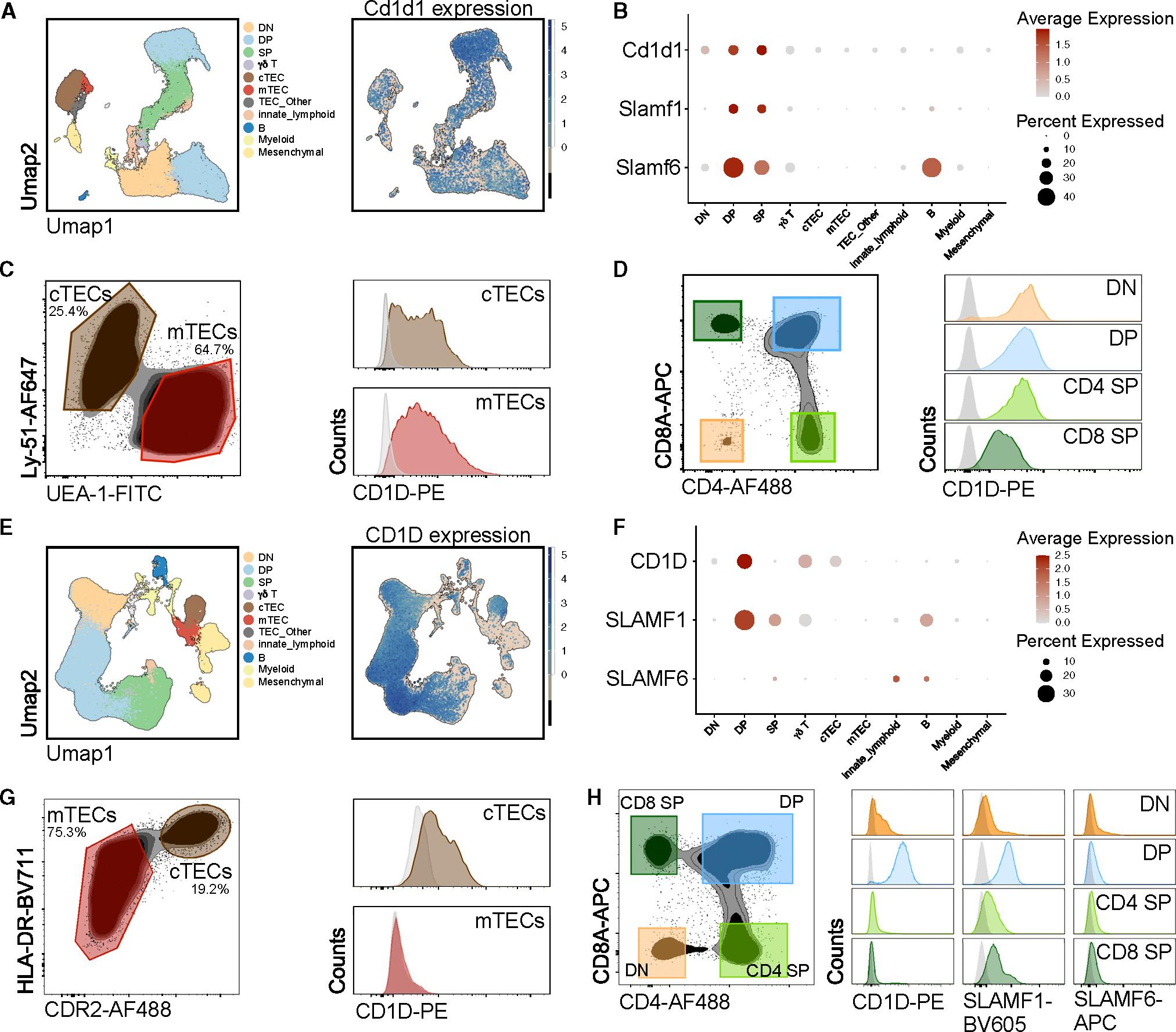
The "innate-like" T cell compartment, known as Tinn, represents a diverse group of T cells that straddle the boundary between innate and adaptive immunity. We explore the transcriptional landscape of Tinn compared to conventional T cells (Tconv) in the human thymus and blood using single-cell RNA sequencing (scRNA-seq) and flow cytometry. In human blood, the majority of Tinn cells share an effector program driven by specific transcription factors, distinct from those governing Tconv cells. Conversely, only a fraction of thymic Tinn cells displays an effector phenotype, while others share transcriptional features with developing Tconv cells, indicating potential divergent developmental pathways. Unlike the mouse, human Tinn cells do not differentiate into multiple effector subsets but develop a mixed type 1/type 17 effector potential. Cross-species analysis uncovers species-specific distinctions, including the absence of type 2 Tinn cells in humans, which implies distinct immune regulatory mechanisms across species.
Keywords: CP: Immunology; MAIT; T cell development; gamma delta T cells; iNKT; thymus.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Update of
-
Unraveling the Phenotypic States of Human innate-like T Cells: Comparative Insights with Conventional T Cells and Mouse Models.bioRxiv [Preprint]. 2023 Dec 8:2023.12.07.570707. doi: 10.1101/2023.12.07.570707. bioRxiv. 2023. Update in: Cell Rep. 2024 Sep 24;43(9):114705. doi: 10.1016/j.celrep.2024.114705. PMID: 38105962 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
