

Alveolin proteins in the Toxoplasma inner membrane complex form a highly interconnected structure that maintains parasite shape and replication
- PMID: 39264987
- PMCID: PMC11421793
- DOI: 10.1371/journal.pbio.3002809
Alveolin proteins in the Toxoplasma inner membrane complex form a highly interconnected structure that maintains parasite shape and replication
Abstract
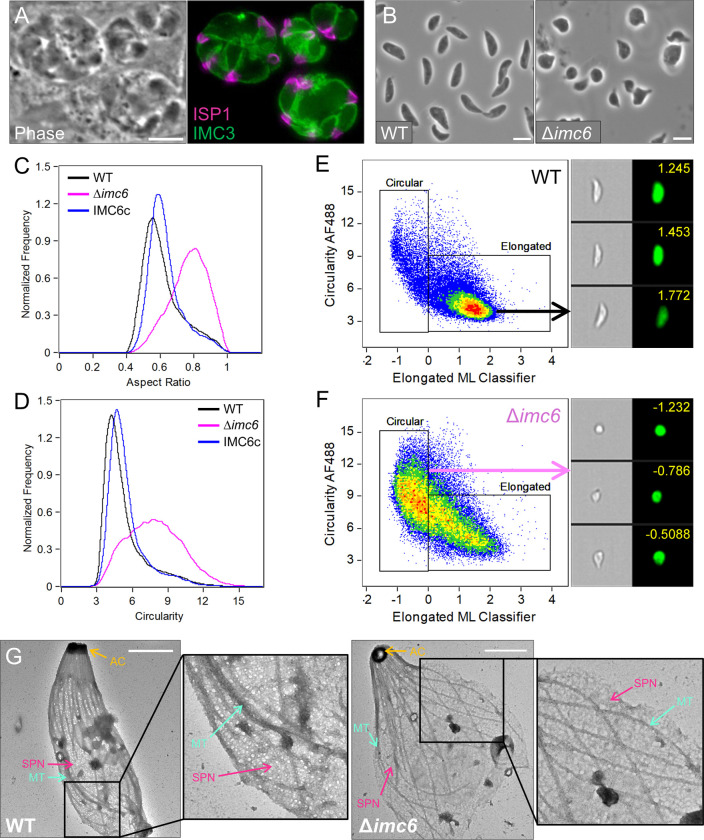
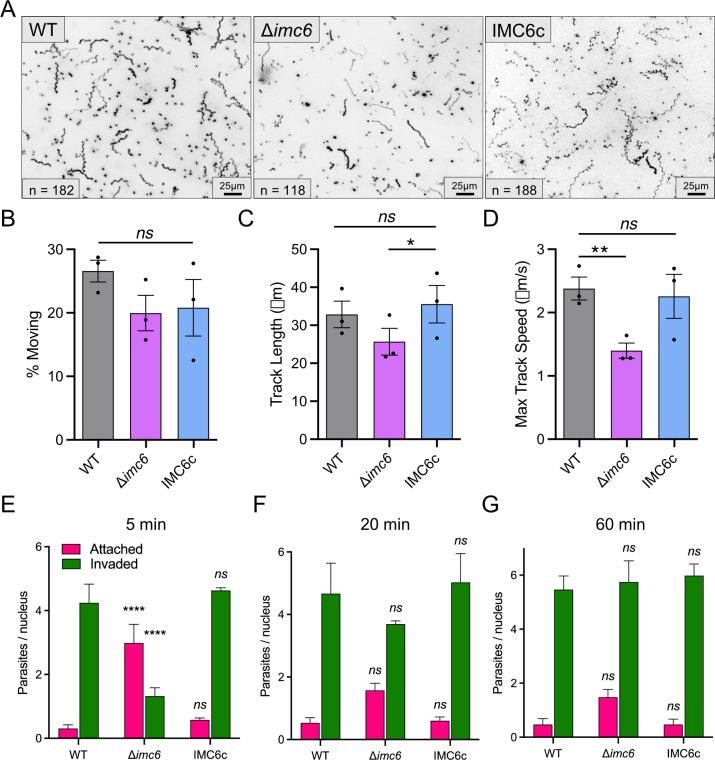
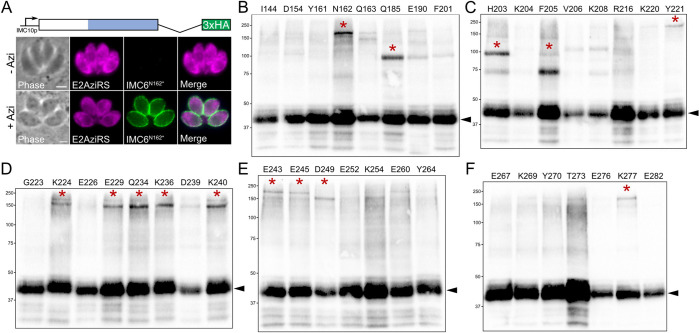
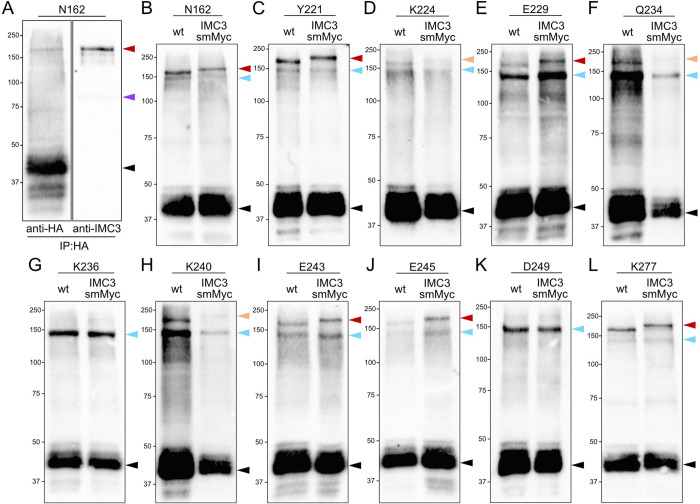
Apicomplexan parasites possess several specialized structures to invade their host cells and replicate successfully. One of these is the inner membrane complex (IMC), a peripheral membrane-cytoskeletal system underneath the plasma membrane. It is composed of a series of flattened, membrane-bound vesicles and a cytoskeletal subpellicular network (SPN) comprised of intermediate filament-like proteins called alveolins. While the alveolin proteins are conserved throughout the Apicomplexa and the broader Alveolata, their precise functions and interactions remain poorly understood. Here, we describe the function of one of these alveolin proteins in Toxoplasma, IMC6. Disruption of IMC6 resulted in striking morphological defects that led to aberrant invasion and replication but surprisingly minor effects on motility. Deletion analyses revealed that the alveolin domain alone is largely sufficient to restore localization and partially sufficient for function. As this highlights the importance of the IMC6 alveolin domain, we implemented unnatural amino acid photoreactive crosslinking to the alveolin domain and identified multiple binding interfaces between IMC6 and 2 other cytoskeletal IMC proteins-IMC3 and ILP1. This provides direct evidence of protein-protein interactions in the alveolin domain and supports the long-held hypothesis that the alveolin domain is responsible for filament formation. Collectively, our study features the conserved alveolin proteins as critical components that maintain the parasite's structural integrity and highlights the alveolin domain as a key mediator of SPN architecture.
Copyright: © 2024 Back et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





Similar articles
-
The Toxoplasma subpellicular network is highly interconnected and defines parasite shape for efficient motility and replication.bioRxiv [Preprint]. 2023 Aug 10:2023.08.10.552545. doi: 10.1101/2023.08.10.552545. bioRxiv. 2023. PMID: 37609316 Free PMC article. Preprint.
-
A photoactivatable crosslinking system reveals protein interactions in the Toxoplasma gondii inner membrane complex.PLoS Biol. 2019 Oct 4;17(10):e3000475. doi: 10.1371/journal.pbio.3000475. eCollection 2019 Oct. PLoS Biol. 2019. PMID: 31584943 Free PMC article.
-
Toxoplasma IMC1 is a central component of the subpellicular network and plays critical roles in parasite morphology, replication, and infectivity.bioRxiv [Preprint]. 2025 Jun 4:2025.06.03.657516. doi: 10.1101/2025.06.03.657516. bioRxiv. 2025. PMID: 40501855 Free PMC article. Preprint.
-
A Novel Toxoplasma Inner Membrane Complex Suture-Associated Protein Regulates Suture Protein Targeting and Colocalizes with Membrane Trafficking Machinery.mBio. 2021 Oct 26;12(5):e0245521. doi: 10.1128/mBio.02455-21. Epub 2021 Oct 12. mBio. 2021. PMID: 34634933 Free PMC article.
-
Microneme proteins: structural and functional requirements to promote adhesion and invasion by the apicomplexan parasite Toxoplasma gondii.Int J Parasitol. 2001 Oct;31(12):1293-302. doi: 10.1016/s0020-7519(01)00257-0. Int J Parasitol. 2001. PMID: 11566297 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
