

Developmental regulation of dermal adipose tissue by BCL11b
- PMID: 39266447
- PMCID: PMC11444185
- DOI: 10.1101/gad.351907.124
Developmental regulation of dermal adipose tissue by BCL11b
Abstract
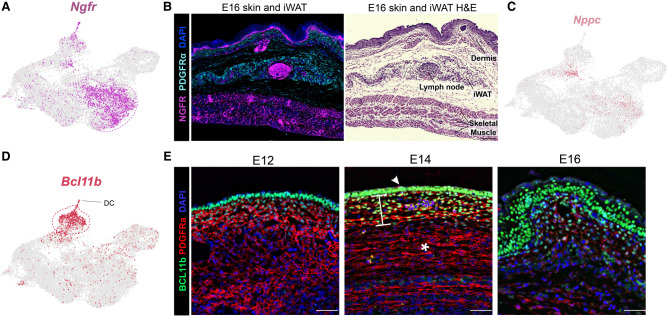
The distinct anatomic environment in which adipose tissues arise during organogenesis is a principle determinant of their adult expansion capacity. Metabolic disease results from a deficiency in hyperplastic adipose expansion within the dermal/subcutaneous depot; thus, understanding the embryonic origins of dermal adipose is imperative. Using single-cell transcriptomics throughout murine embryogenesis, we characterized cell populations, including Bcl11b + cells, that regulate the development of dermal white adipose tissue (dWAT). We discovered that BCL11b expression modulates the Wnt signaling microenvironment to enable adipogenic differentiation in the dermal compartment. Subcutaneous and visceral adipose arises from a distinct population of Nefl + cells during embryonic organogenesis, whereas Pi16 + /Dpp4 + fibroadipogenic progenitors support obesity-stimulated hypertrophic expansion in the adult. Together, these results highlight the unique regulatory pathways used by anatomically distinct adipose depots, with important implications for human metabolic disease.
Keywords: Bcl11b; FAP; Nefl; Ngfr; Pi16; adipose development; adipose lineage; dermal adipose; single cell.
© 2024 Traynor et al.; Published by Cold Spring Harbor Laboratory Press.
Figures





References
-
- Belligoli A, Compagnin C, Sanna M, Favaretto F, Fabris R, Busetto L, Foletto M, Dal Prà C, Serra R, Prevedello L, et al. 2019. Characterization of subcutaneous and omental adipose tissue in patients with obesity and with different degrees of glucose impairment. Sci Rep 9: 11333. 10.1038/s41598-019-47719-y - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous