

Exploiting human immune repertoire transgenic mice for protective monoclonal antibodies against antimicrobial resistant Acinetobacter baumannii
- PMID: 39266557
- PMCID: PMC11392949
- DOI: 10.1038/s41467-024-52357-8
Exploiting human immune repertoire transgenic mice for protective monoclonal antibodies against antimicrobial resistant Acinetobacter baumannii
Abstract
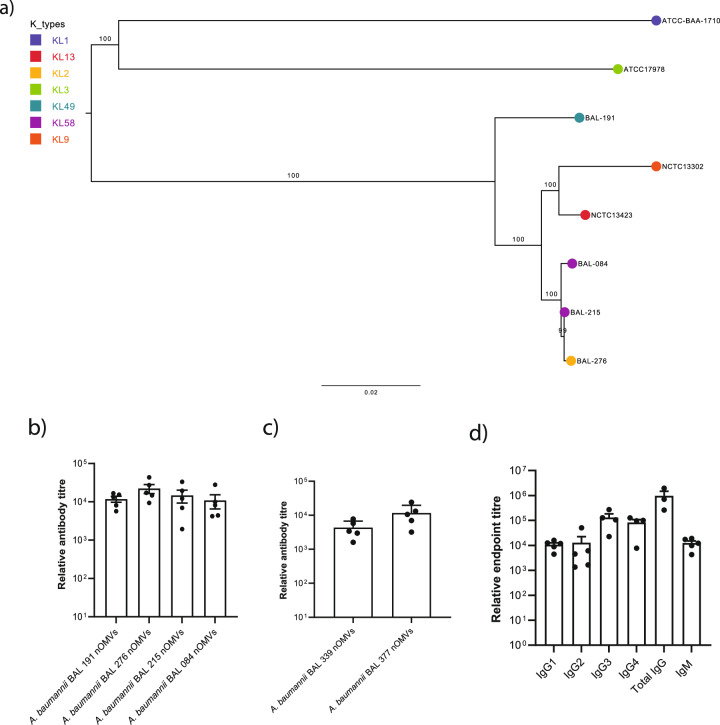
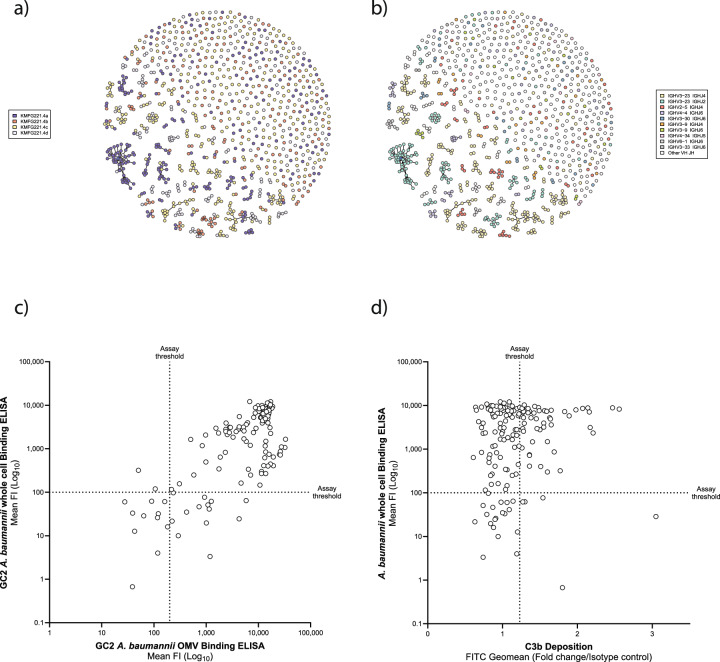
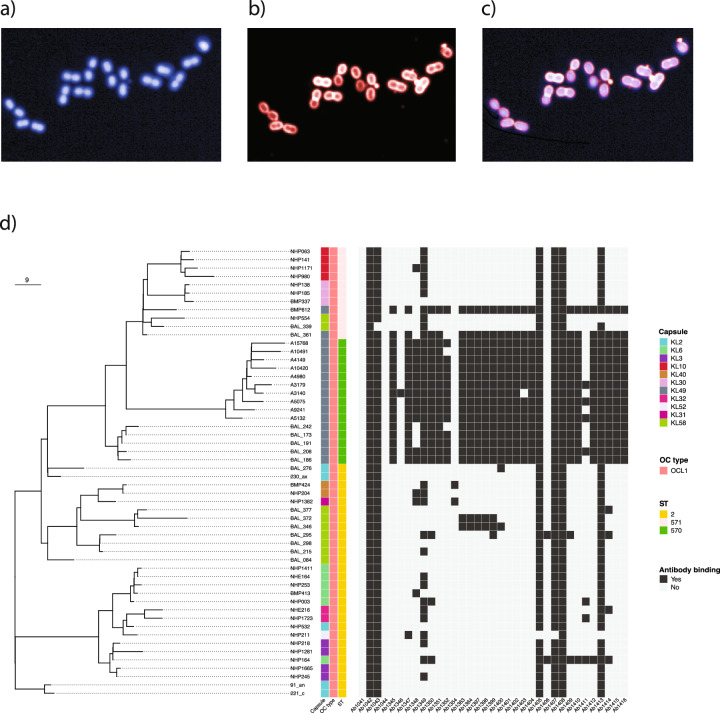
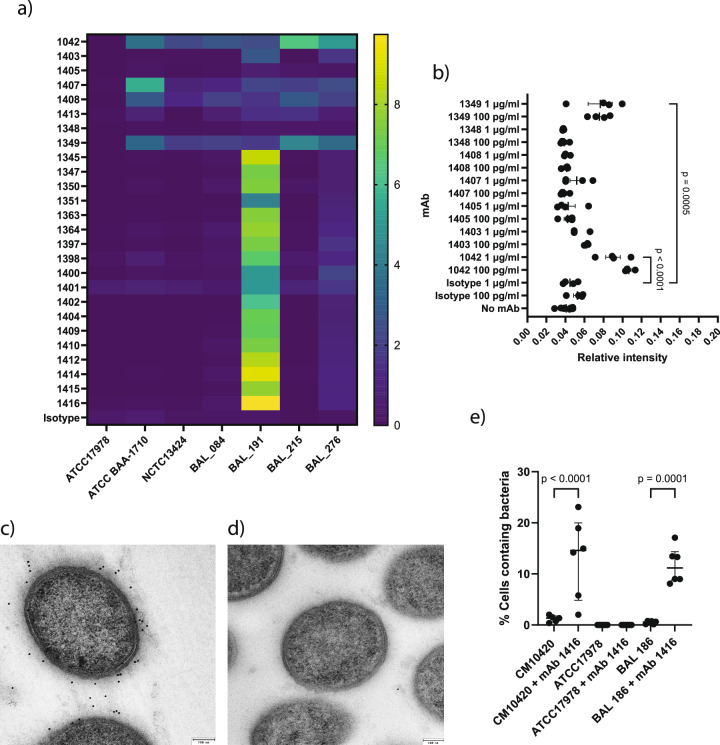
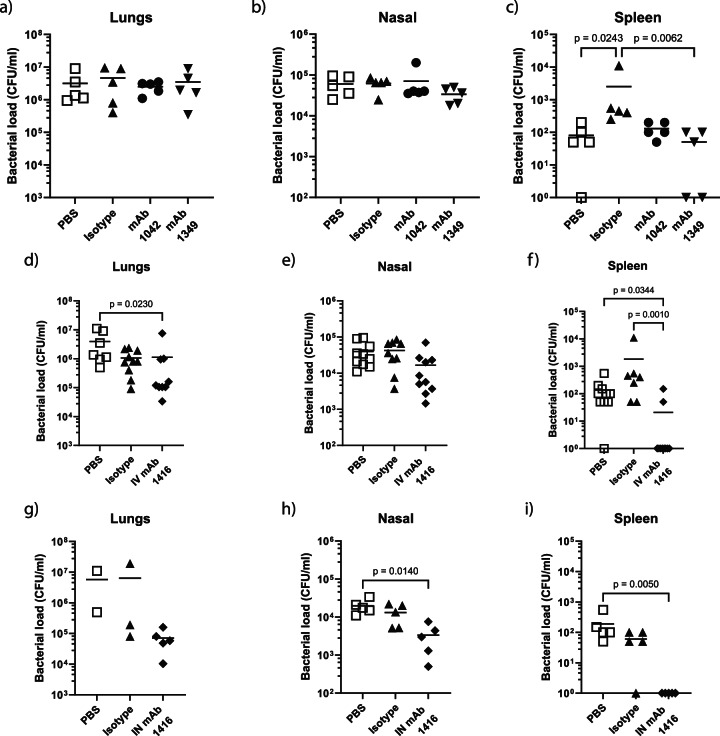
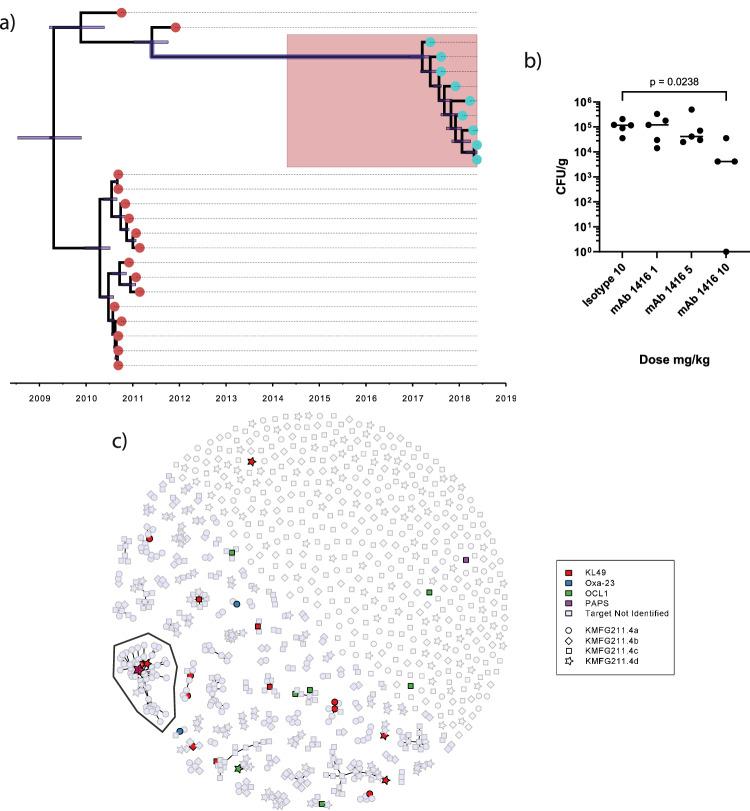
The use of monoclonal antibodies for the control of drug resistant nosocomial bacteria may alleviate a reliance on broad spectrum antimicrobials for treatment of infection. We identify monoclonal antibodies that may prevent infection caused by carbapenem resistant Acinetobacter baumannii. We use human immune repertoire mice (Kymouse platform mice) as a surrogate for human B cell interrogation to establish an unbiased strategy to probe the antibody-accessible target landscape of clinically relevant A. baumannii. After immunisation of the Kymouse platform mice with A. baumannii derived outer membrane vesicles (OMV) we identify 297 antibodies and analyse 26 of these for functional potential. These antibodies target lipooligosaccharide (OCL1), the Oxa-23 protein, and the KL49 capsular polysaccharide. We identify a single monoclonal antibody (mAb1416) recognising KL49 capsular polysaccharide to demonstrate prophylactic in vivo protection against a carbapenem resistant A. baumannii lineage associated with neonatal sepsis mortality in Asia. Our end-to-end approach identifies functional monoclonal antibodies with prophylactic potential against major lineages of drug resistant bacteria accounting for phylogenetic diversity and clinical relevance without existing knowledge of a specific target antigen. Such an approach might be scaled for a additional clinically important bacterial pathogens in the post-antimicrobial era.
© 2024. The Author(s).
Conflict of interest statement
The authors declare the following competing interests. J.B.S., Q.Z., and S.T.R. are employees of Kymab Ltd, a Sanofi company and may have held or continue to hold stock options or shares in Sanofi. S.O’.L., J.B.S., A.K., S.W., S.T.R., and S.B. are listed as inventors on a published patent WO2023094628A1 covering monoclonal antibodies generated by this work. S.O’.L., A.K., I.A., B.S., H.C., V.W., N.F.O., D.M.S., C.P., S.W., and P.K. were employees of Kymab, a Sanofi Company within the last three years and may have held or continue to hold stock options or shares in Sanofi. The other authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
