

The subcellular distribution of miRNA isoforms, tRNA-derived fragments, and rRNA-derived fragments depends on nucleotide sequence and cell type
- PMID: 39267057
- PMCID: PMC11397057
- DOI: 10.1186/s12915-024-01970-6
The subcellular distribution of miRNA isoforms, tRNA-derived fragments, and rRNA-derived fragments depends on nucleotide sequence and cell type
Abstract
Background: MicroRNA isoforms (isomiRs), tRNA-derived fragments (tRFs), and rRNA-derived fragments (rRFs) represent most of the small non-coding RNAs (sncRNAs) found in cells. Members of these three classes modulate messenger RNA (mRNA) and protein abundance and are dysregulated in diseases. Experimental studies to date have assumed that the subcellular distribution of these molecules is well-understood, independent of cell type, and the same for all isoforms of a sncRNA.
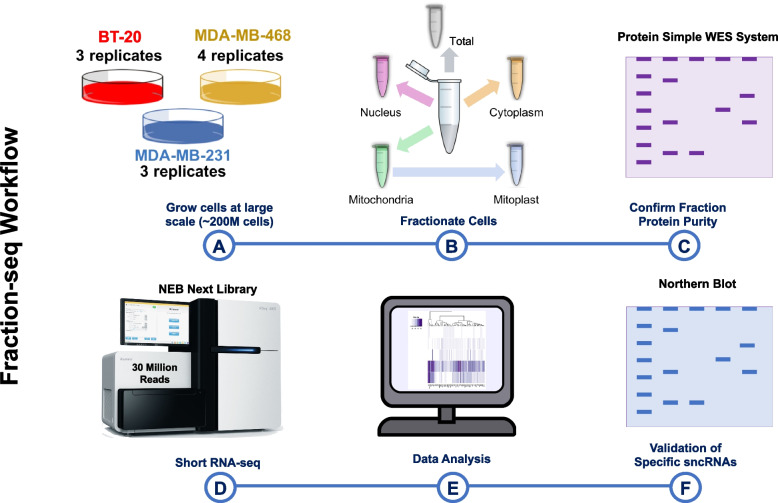
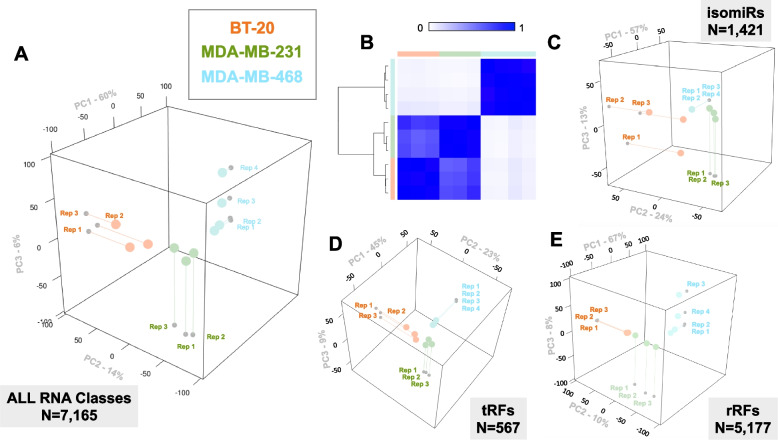
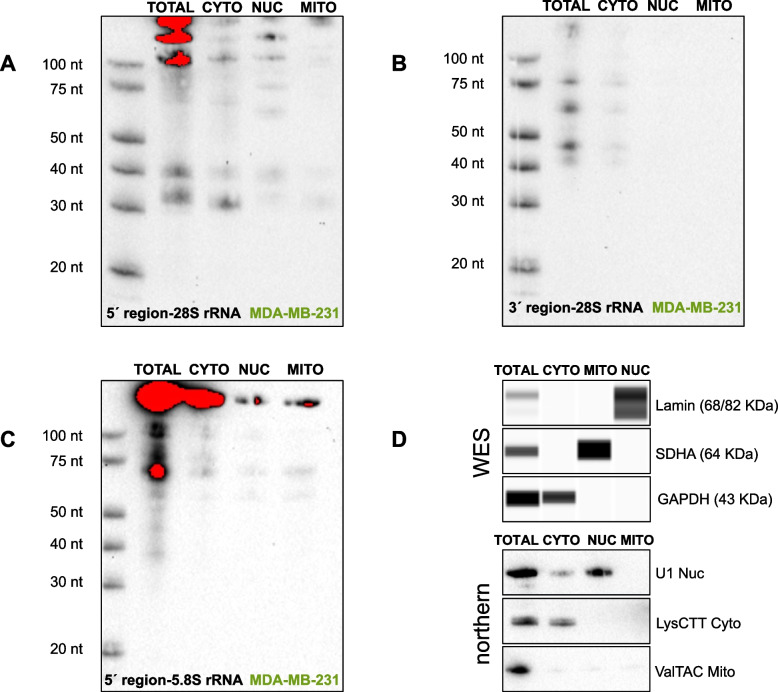
Results: We tested these assumptions by investigating the subcellular distribution of isomiRs, tRFs, and rRFs in biological replicates from three cell lines from the same tissue and same-sex donors that model the same cancer subtype. In each cell line, we profiled the isomiRs, tRFs, and rRFs in the nucleus, cytoplasm, whole mitochondrion (MT), mitoplast (MP), and whole cell. Using a rigorous mathematical model we developed, we accounted for cross-fraction contamination and technical errors and adjusted the measured abundances accordingly. Analyses of the adjusted abundances show that isomiRs, tRFs, and rRFs exhibit complex patterns of subcellular distributions. These patterns depend on each sncRNA's exact sequence and the cell type. Even in the same cell line, isoforms of the same sncRNA whose sequences differ by a few nucleotides (nts) can have different subcellular distributions.
Conclusions: SncRNAs with similar sequences have different subcellular distributions within and across cell lines, suggesting that each isoform could have a different function. Future computational and experimental studies of isomiRs, tRFs, and rRFs will need to distinguish among each molecule's various isoforms and account for differences in each isoform's subcellular distribution in the cell line at hand. While the findings add to a growing body of evidence that isomiRs, tRFs, rRFs, tRNAs, and rRNAs follow complex intracellular trafficking rules, further investigation is needed to exclude alternative explanations for the observed subcellular distribution of sncRNAs.
Keywords: Small non-coding RNAs; isomiRs; miRNA isoforms; miRNAs; microRNAs; rRFs; rRNA-derived fragments; sncRNAs; subcellular distribution; tRFs; tRNA-derived fragments.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
