

Discovery of vitexin as a novel VDR agonist that mitigates the transition from chronic intestinal inflammation to colorectal cancer
- PMID: 39272040
- PMCID: PMC11395219
- DOI: 10.1186/s12943-024-02108-6
Discovery of vitexin as a novel VDR agonist that mitigates the transition from chronic intestinal inflammation to colorectal cancer
Abstract
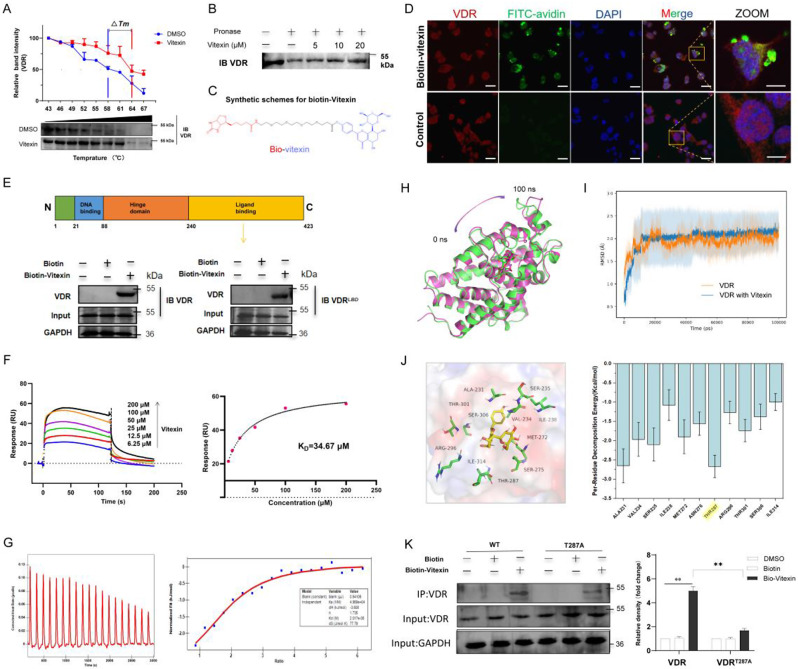
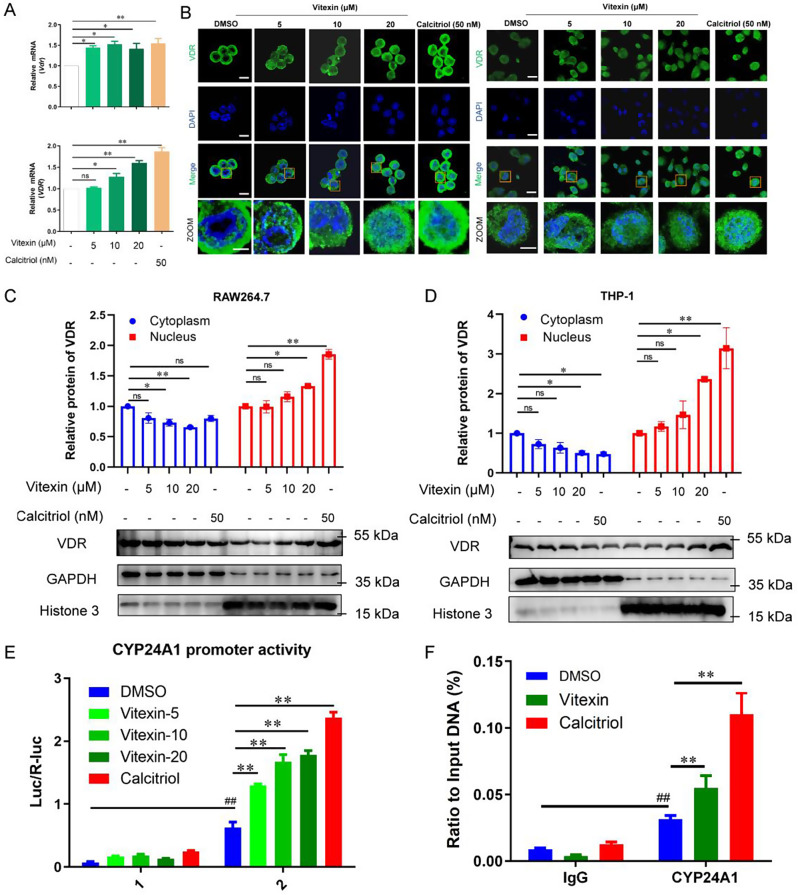
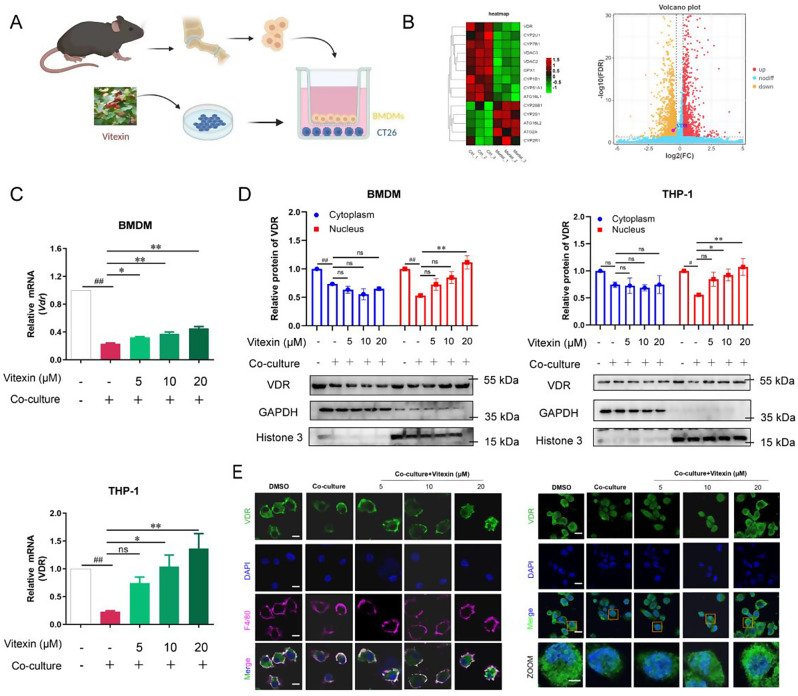
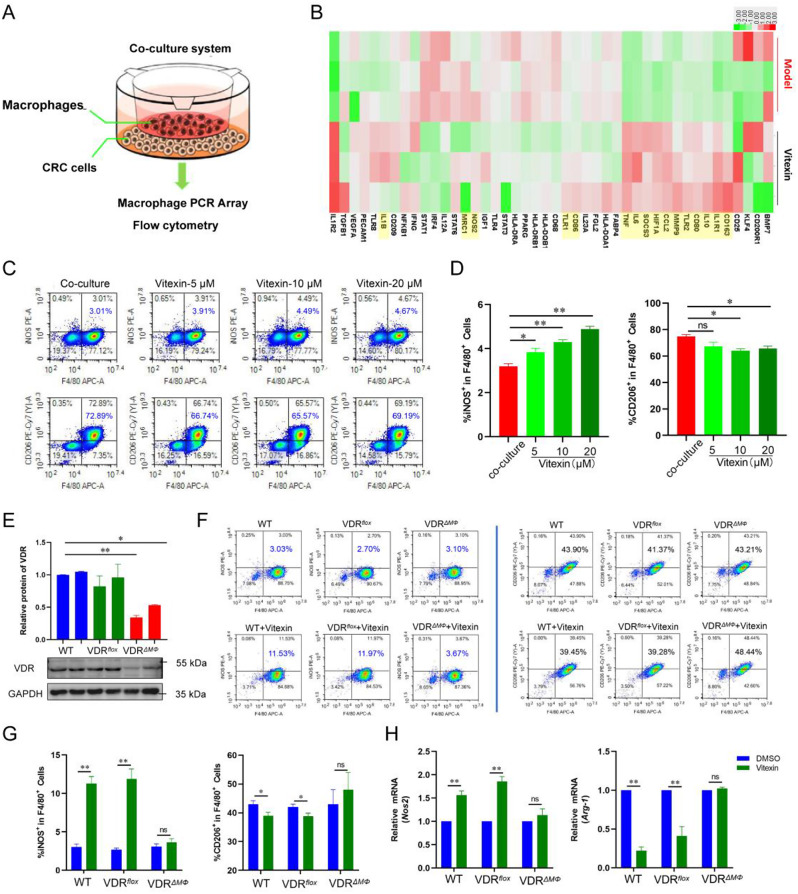
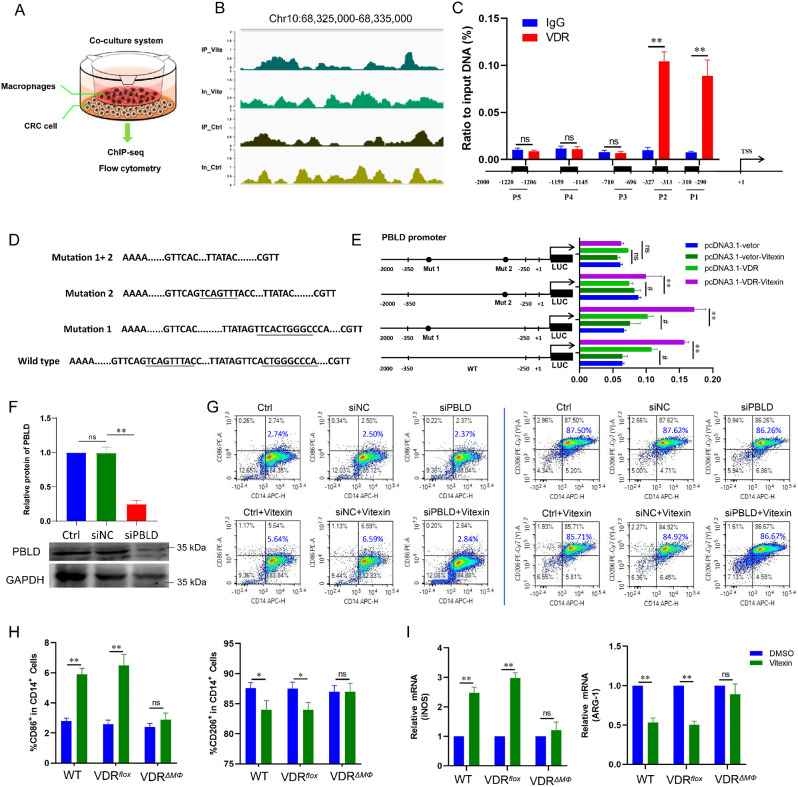
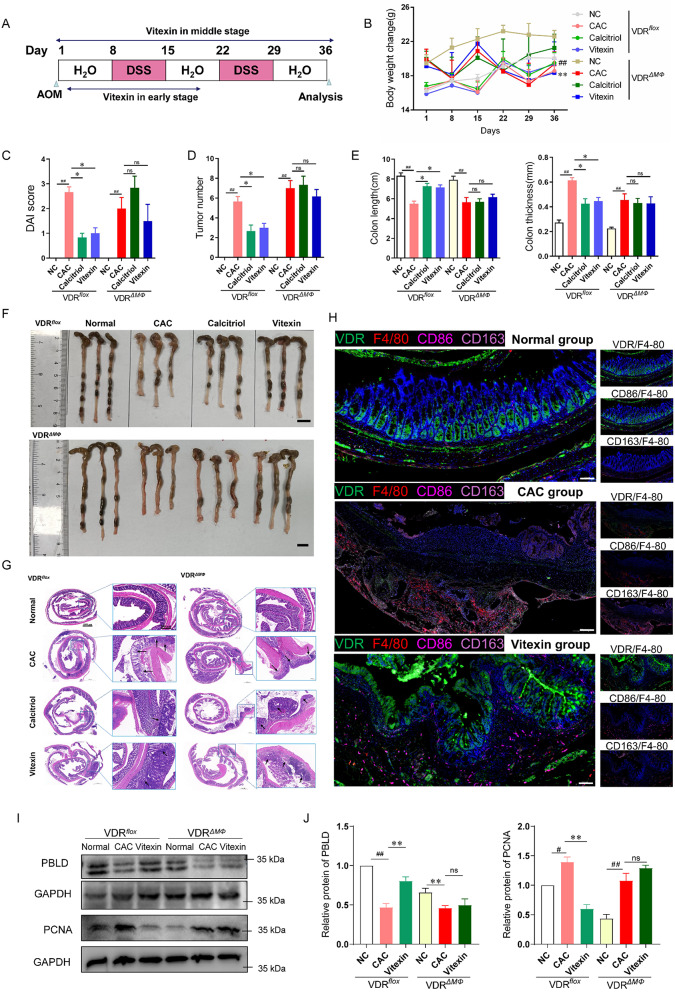
Colitis-associated colorectal cancer (CAC) frequently develops in patients with inflammatory bowel disease (IBD) who have been exposed to a prolonged state of chronic inflammation. The investigation of pharmacological agents and their mechanisms to prevent precancerous lesions and inhibit their progression remains a significant focus and challenge in CAC research. Previous studies have demonstrated that vitexin effectively mitigates CAC, however, its precise mechanism of action warrants further exploration. This study reveals that the absence of the Vitamin D receptor (VDR) accelerates the progression from chronic colitis to colorectal cancer. Our findings indicate that vitexin can specifically target the VDR protein, facilitating its translocation into the cell nucleus to exert transcriptional activity. Additionally, through a co-culture model of macrophages and cancer cells, we observed that vitexin promotes the polarization of macrophages towards the M1 phenotype, a process that is dependent on VDR. Furthermore, ChIP-seq analysis revealed that vitexin regulates the transcriptional activation of phenazine biosynthesis-like domain protein (PBLD) via VDR. ChIP assays and dual luciferase reporter assays were employed to identify the functional PBLD regulatory region, confirming that the VDR/PBLD pathway is critical for vitexin-mediated regulation of macrophage polarization. Finally, in a mouse model with myeloid VDR gene knockout, we found that the protective effects of vitexin were abolished in mid-stage CAC. In summary, our study establishes that vitexin targets VDR and modulates macrophage polarization through the VDR/PBLD pathway, thereby alleviating the transition from chronic colitis to colorectal cancer.
Keywords: Colitis-associated colorectal cancer; Macrophage; Vitamin D receptor; Vitexin.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Vitamin D receptor regulates methyltransferase like 14 to mitigate colitis-associated colorectal cancer.J Genet Genomics. 2025 Aug;52(8):1011-1020. doi: 10.1016/j.jgg.2024.12.020. Epub 2025 Jan 6. J Genet Genomics. 2025. PMID: 39778713
-
VDR promotes pancreatic cancer progression in vivo by activating CCL20-mediated M2 polarization of tumor associated macrophage.Cell Commun Signal. 2024 Apr 10;22(1):224. doi: 10.1186/s12964-024-01578-x. Cell Commun Signal. 2024. PMID: 38600588 Free PMC article.
-
Weissella cibaria suppresses colitis-associated colorectal cancer by modulating the gut microbiota-bile acid-FXR axis.mSystems. 2025 Jul 22;10(7):e0028825. doi: 10.1128/msystems.00288-25. Epub 2025 Jul 3. mSystems. 2025. PMID: 40607831 Free PMC article.
-
Strategies for detecting colon cancer and/or dysplasia in patients with inflammatory bowel disease.Cochrane Database Syst Rev. 2006 Apr 19;(2):CD000279. doi: 10.1002/14651858.CD000279.pub3. Cochrane Database Syst Rev. 2006. Update in: Cochrane Database Syst Rev. 2017 Sep 18;9:CD000279. doi: 10.1002/14651858.CD000279.pub4. PMID: 16625534 Updated.
-
Astragaloside IV: A Promising Drug to Prevent the Transition from Colitis to Colorectal Cancer.Am J Chin Med. 2025;53(4):1065-1091. doi: 10.1142/S0192415X25500405. Epub 2025 Jun 18. Am J Chin Med. 2025. PMID: 40530759 Review.
Cited by
-
Dietary Flavonoids Vitexin and Isovitexin: New Insights into Their Functional Roles in Human Health and Disease Prevention.Int J Mol Sci. 2025 Jul 21;26(14):6997. doi: 10.3390/ijms26146997. Int J Mol Sci. 2025. PMID: 40725244 Free PMC article. Review.
-
Monotropein inhibits epithelial-mesenchymal transition in chronic colitis via the mTOR/P70S6K pathway.Front Pharmacol. 2025 Feb 18;16:1536091. doi: 10.3389/fphar.2025.1536091. eCollection 2025. Front Pharmacol. 2025. PMID: 40041493 Free PMC article.
-
Honokiol-Magnolol-Baicalin Possesses Synergistic Anticancer Potential and Enhances the Efficacy of Anti-PD-1 Immunotherapy in Colorectal Cancer by Triggering GSDME-Dependent Pyroptosis.Adv Sci (Weinh). 2025 Apr;12(13):e2417022. doi: 10.1002/advs.202417022. Epub 2025 Feb 14. Adv Sci (Weinh). 2025. PMID: 39950759 Free PMC article.
-
Detecting PI3K and TP53 Pathway Disruptions in Early-Onset Colorectal Cancer Among Hispanic/Latino Patients.Cancer Med. 2025 Apr;14(7):e70791. doi: 10.1002/cam4.70791. Cancer Med. 2025. PMID: 40165548 Free PMC article.
-
Deuterium-depleted water inhibits the malignant progression of colorectal cancer cells by modulating oxidative stress.Oncol Rep. 2025 Jun;53(6):70. doi: 10.3892/or.2025.8903. Epub 2025 May 2. Oncol Rep. 2025. PMID: 40314087 Free PMC article.
References
-
- Fujita M, Matsubara N, Matsuda I, Maejima K, Oosawa A, Yamano T, Fujimoto A, Furuta M, Nakano K, Oku-Sasaki A, et al. Genomic landscape of colitis-associated cancer indicates the impact of chronic inflammation and its stratification by mutations in the wnt signaling. Oncotarget. 2018;9:969–81. 10.18632/oncotarget.22867 - DOI - PMC - PubMed
-
- Xu Y, Liu K, Li C, Li M, Liu F, Zhou X, Sun M, Ranganathan M, Zhang L, Wang S et al. The largest Chinese cohort study indicates homologous recombination pathway gene mutations as another major genetic risk factor for colorectal Cancer with heterogeneous clinical phenotypes. Research 2023, 6. - PMC - PubMed
-
- Yaeger R, Shah MA, Miller VA, Kelsen JR, Wang K, Heins ZJ, Ross JS, He Y, Sanford E, Yantiss RK, et al. Genomic alterations observed in Colitis-Associated Cancers are distinct from those found in sporadic colorectal cancers and vary by type of inflammatory bowel disease. Gastroenterology. 2016;151:278–. 10.1053/j.gastro.2016.04.001 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
