

Stabilization of dimeric PYR/PYL/RCAR family members relieves abscisic acid-induced inhibition of seed germination
- PMID: 39277642
- PMCID: PMC11401921
- DOI: 10.1038/s41467-024-52426-y
Stabilization of dimeric PYR/PYL/RCAR family members relieves abscisic acid-induced inhibition of seed germination
Abstract
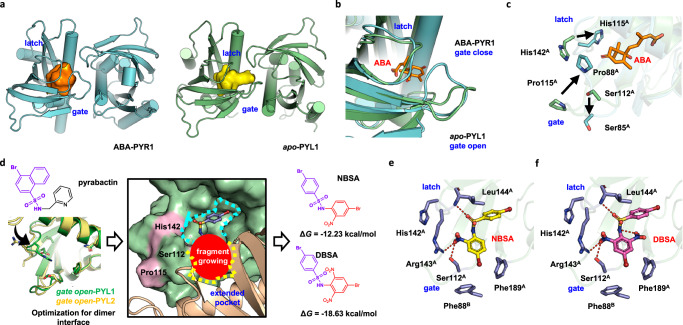
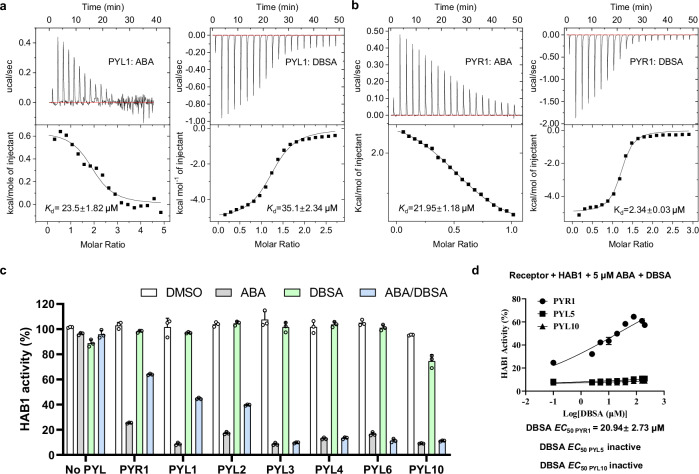
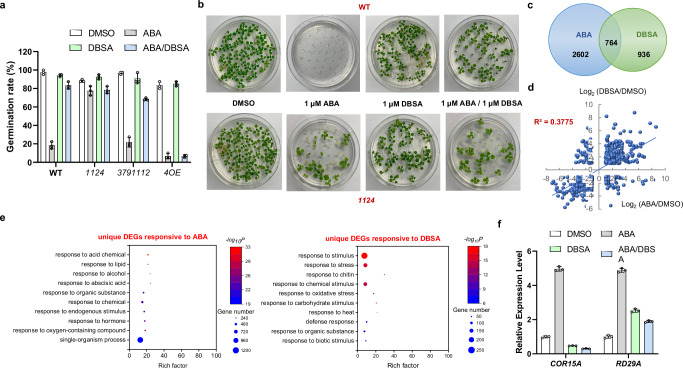
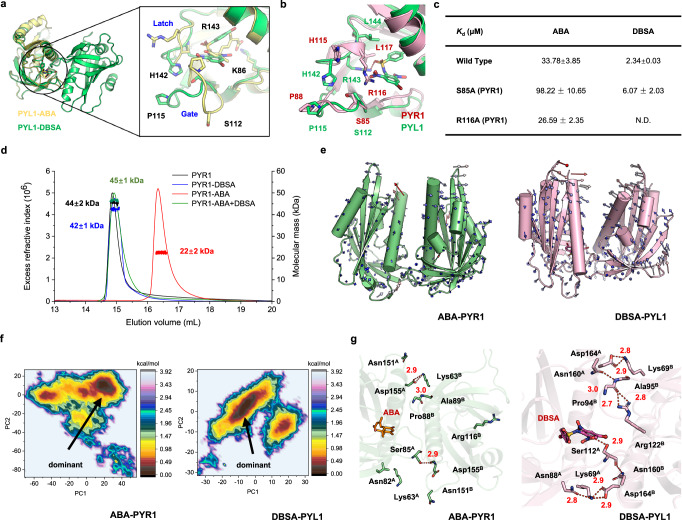
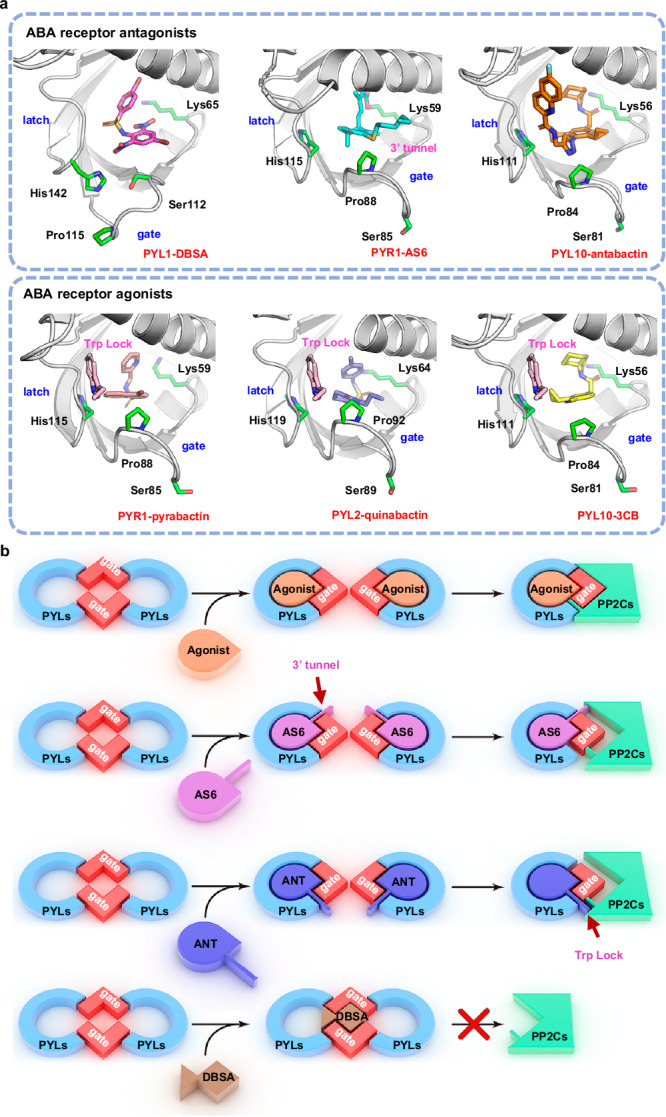
Abscisic acid (ABA) is the primary preventing factor of seed germination, which is crucial to plant survival and propagation. ABA-induced seed germination inhibition is mainly mediated by the dimeric PYR/PYL/RCAR (PYLs) family members. However, little is known about the relevance between dimeric stability of PYLs and seed germination. Here, we reveal that stabilization of PYL dimer can relieve ABA-induced inhibition of seed germination using chemical genetic approaches. Di-nitrobensulfamide (DBSA), a computationally designed chemical probe, yields around ten-fold improvement in receptor affinity relative to ABA. DBSA reverses ABA-induced inhibition of seed germination mainly through dimeric receptors and recovers the expression of ABA-responsive genes. DBSA maintains PYR1 in dimeric state during protein oligomeric state experiment. X-ray crystallography shows that DBSA targets a pocket in PYL dimer interface and may stabilize PYL dimer by forming hydrogen networks. Our results illustrate the potential of PYL dimer stabilization in preventing ABA-induced seed germination inhibition.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Challinor, A. J., Koehler, A. K., Ramirez-Villegas, J., Whitfield, S. & Das, B. Current warming will reduce yields unless maize breeding and seed systems adapt immediately. Nat. Clim. Change6, 954–958 (2016). 10.1038/nclimate3061 - DOI
-
- Miransari, M. & Smith, D. L. Plant hormones and seed germination. Environ. Exp. Bot.99, 110–121 (2014). 10.1016/j.envexpbot.2013.11.005 - DOI
Publication types
MeSH terms
Substances
Associated data
- BioProject/PRJNA1138801
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
