

Mitonuclear and phenotypic discordance in an Atlantic Forest frog hybrid zone
- PMID: 39279790
- PMCID: PMC11393776
- DOI: 10.1002/ece3.70262
Mitonuclear and phenotypic discordance in an Atlantic Forest frog hybrid zone
Abstract
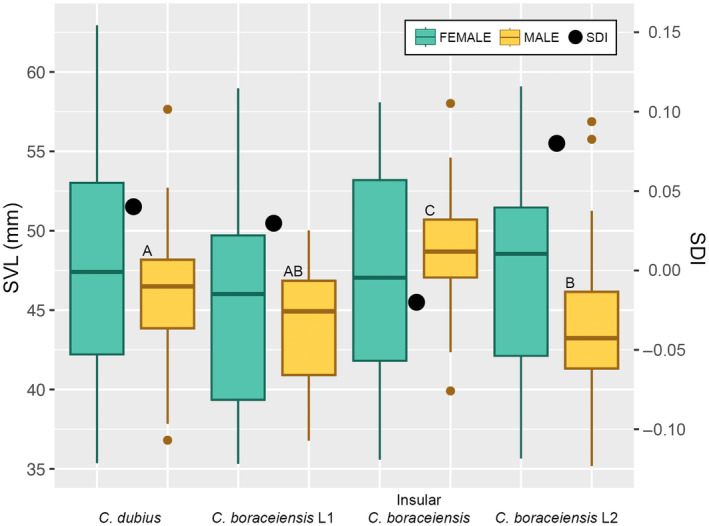
Discordance between mitochondrial and nuclear DNA is common among animals and can be the result of a number of evolutionary processes, including incomplete lineage sorting and introgression. Particularly relevant in contact zones, mitonuclear discordance is expected because the mitochondrial genome is haploid and primarily uniparentally inherited, whereas nuclear loci are evolving at slower rates. In addition, when closely related taxa come together in hybrid zones, the distribution of diagnostic phenotypic characters and their concordance with the mitochondrial or nuclear lineages can also inform on historical and ongoing dynamics within hybrid zones. Overall, genetic and phenotypic discordances provide evidence for evolutionary divergence and processes that maintain boundaries among sister species or lineages. In this study, we characterized patterns of genetic and phenotypic variation in a contact zone between Cycloramphus dubius and Cycloramphus boraceiensis, two sister species of frogs endemic to the Atlantic Coastal Forest of Brazil. We examined genomic-scale nuclear diversification across 19 populations, encompassing the two parental forms and a contact zone between them. We compared the distribution of genomic DNA variability with that of a mitochondrial locus (16S) and two morphological traits (dorsal tubercles and body size). Our results reveal multiple divergent lineages with ongoing admixture. We detected discordance in patterns of introgression across the three data types. Cycloramphus dubius males are significantly larger than C. boraceiensis males, and we posit that competition among males in the hybrid zone, coupled with mate choice by females, may be one mechanism leading to patterns of introgression observed between the species.
Keywords: anuran; body size; intrasexual competition; introgression; mitonuclear discordance; sexual selection.
© 2024 The Author(s). Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that there are no conflicts of interest.
Figures






References
-
- Akopyan, M. , Gompert, Z. , Klonoski, K. , Vega, A. , Kaiser, K. , Mackelprang, R. , Rosenblum, E. B. , & Robertson, J. M. (2020). Genetic and phenotypic evidence of a contact zone between divergent colour morphs of the iconic red‐eyed treefrog. Molecular Ecology, 29(22), 4442–4456. 10.1111/mec.15639 - DOI - PubMed
-
- Amaral, F. R. , Alvarado‐Serrano, D. F. , Maldonado‐Coelho, M. , Pellegrino, K. C. M. , Miyaki, C. Y. , Montesanti, J. A. C. , Lima‐Ribeiro, M. S. , Hickerson, M. J. , & Thom, G. (2018). Climate explains recent population divergence, introgression, and persistence in tropical mountains: Phylogenomic evidence from Atlantic Forest warbling finches. bioRxiv, 2018, 439265. 10.1101/439265. - DOI
-
- Amaro, R. C. , Rodrigues, M. T. , Yonenaga‐Yassuda, Y. , & Carnaval, A. C. (2012). Demographic processes in the montane Atlantic rainforest: Molecular and cytogenetic evidence from the endemic frog Proceratophrys boiei . Molecular Phylogenetics and Evolution, 62(3), 880–888. 10.1016/j.ympev.2011.11.004 - DOI - PubMed
-
- Anderson, E. (1948). Hybridization of the habitat. Evolution, 2(1), 1–9. 10.2307/2405610 - DOI
Associated data
LinkOut - more resources
Full Text Sources
