

Adaptive radiation of the Callicarpa genus in the Bonin Islands revealed through double-digest restriction site-associated DNA sequencing analysis
- PMID: 39279792
- PMCID: PMC11393766
- DOI: 10.1002/ece3.70216
Adaptive radiation of the Callicarpa genus in the Bonin Islands revealed through double-digest restriction site-associated DNA sequencing analysis
Abstract
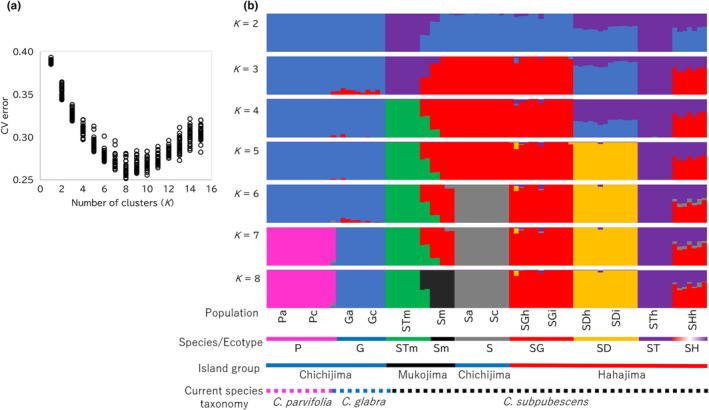
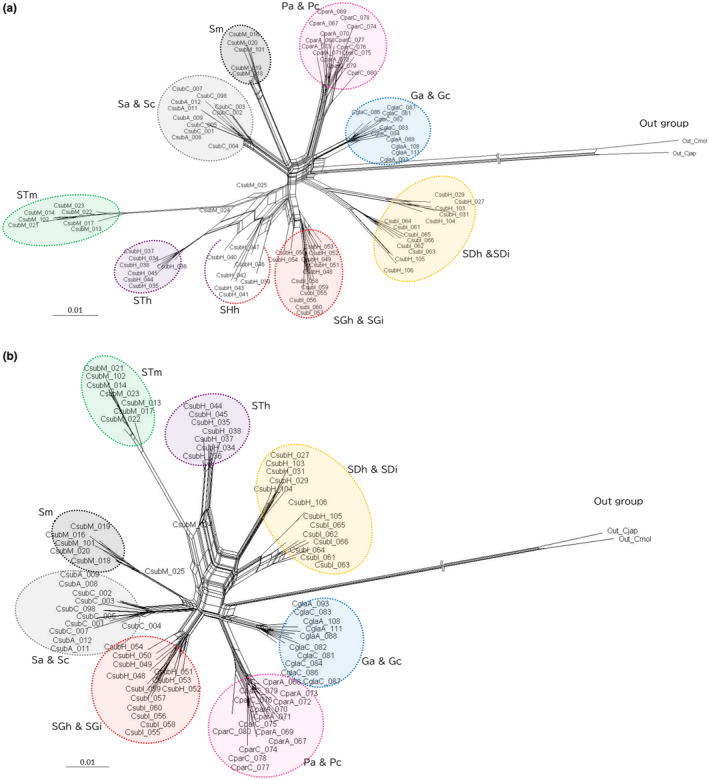
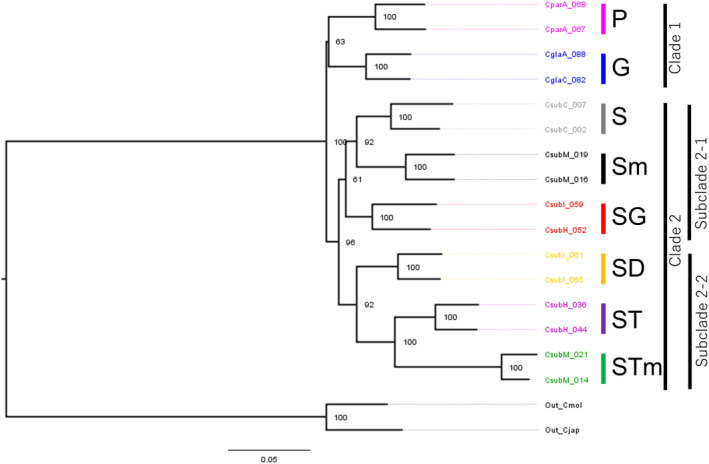
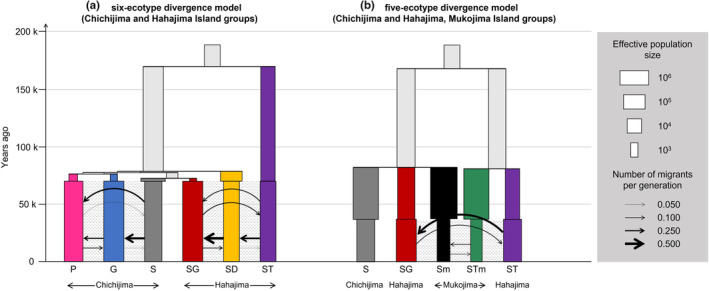
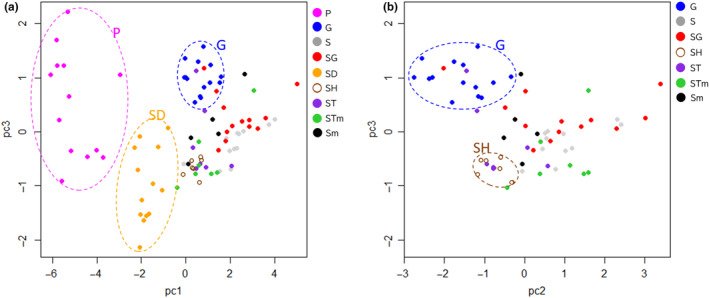
The Bonin Islands, comprised of the Mukojima, Chichijima, and Hahajima Islands, are known for their isolated and distinctive habitats, hosting a diverse array of endemic flora and fauna. In these islands, adaptive radiation has played a remarkable role in speciation, particularly evident in the Callicarpa genus that is represented by three species: Callicarpa parvifolia and C. glabra exclusive to the Chichijima Islands, and Callicarpa subpubescens, distributed across the entire Bonin Islands. Notably, C. subpubescens exhibits multiple ecotypes, differing in leaf hair density, flowering time, and tree size. In this study, we aimed to investigate species and ecotype diversification patterns, estimate divergence times, and explore cryptic species within Callicarpa in the Bonin Islands using phenotypic and genetic data (double-digest restriction site-associated DNA sequencing). Genetic analysis revealed that C. parvifolia and C. glabra both formed single, distinct genetic groups. Conversely, C. subpubescens consisted of six genetic groups corresponding to different ecotypes and regions, and a hybrid group resulting from the hybridization between two of these genetic groups. Population demography analysis focusing on six Chichijima and Hahajima Islands-based species/ecotypes indicated that all species and ecotypes except one ecotype diverged simultaneously around 73-77 kya. The star-shaped neighbor-net tree also suggests the simultaneous divergence of species and ecotypes. The species and ecotypes that simultaneously diverged adapted to dry environments and understory forests, suggesting that aridification may have contributed to this process of adaptive radiation. Moreover, leaf morphology, flowering time, and genetic analyses suggested the presence of two cryptic species and one hybrid species within C. subpubescens.
Keywords: Lamiaceae; Ogasawara Islands; ddRAD‐Seq; oceanic islands; quaternary.
© 2024 The Author(s). Ecology and Evolution published by John Wiley & Sons Ltd.
Conflict of interest statement
Authors declare no competing interests.
Figures
References
-
- Abobatta, W. F. (2021). Fruit orchards under climate change conditions: Adaptation strategies and management. Journal of Applied Biotechnology & Bioengineering, 8(3), 99–102. 10.15406/jabb.2021.08.00260 - DOI
-
- Aedo, C. , Medina, L. , & Fernández‐Albert, M. (2013). Species richness and endemicity in the Spanish vascular flora. Nordic Journal of Botany, 31(4), 478–488. 10.1111/j.1756-1051.2012.00009.x - DOI
-
- Ando, H. , Setsuko, S. , Horikoshi, K. , Suzuki, H. , Umehara, S. , Yamasaki, M. , Hanya, G. , Inoue‐Murayama, M. , & Isagi, Y. (2016). Seasonal and inter‐Island variation in the foraging strategy of the critically endangered red‐headed Wood pigeon Columba janthina nitens in disturbed Island habitats derived from high‐throughput sequencing. Ibis, 158(2), 291–304.
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
