

This is a preprint.
USP37 prevents premature disassembly of stressed replisomes by TRAIP
- PMID: 39282314
- PMCID: PMC11398331
- DOI: 10.1101/2024.09.03.611025
USP37 prevents premature disassembly of stressed replisomes by TRAIP
Update in
-
USP37 prevents premature disassembly of stressed replisomes by TRAIP.Nat Commun. 2025 Jun 18;16(1):5333. doi: 10.1038/s41467-025-60139-z. Nat Commun. 2025. PMID: 40533495 Free PMC article.
Abstract
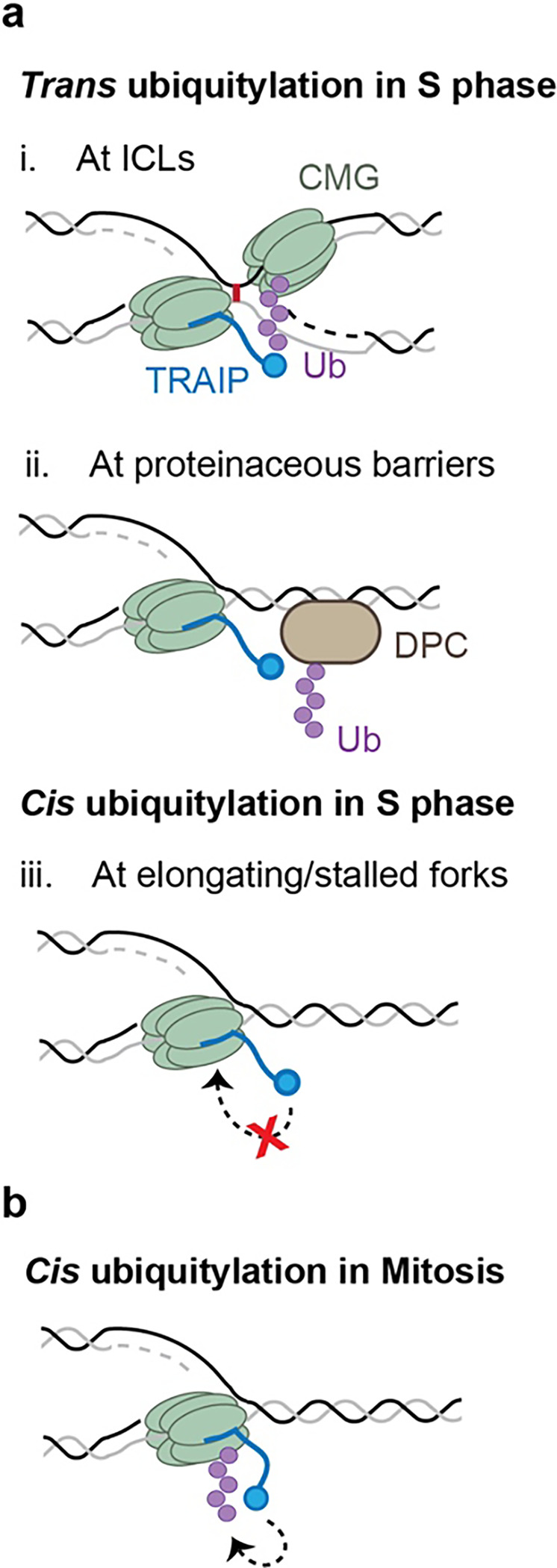
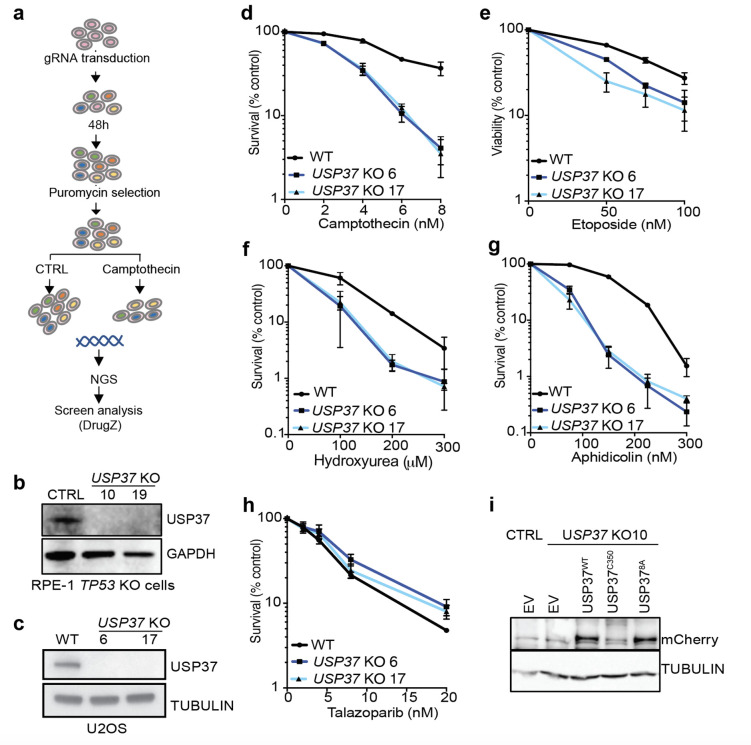
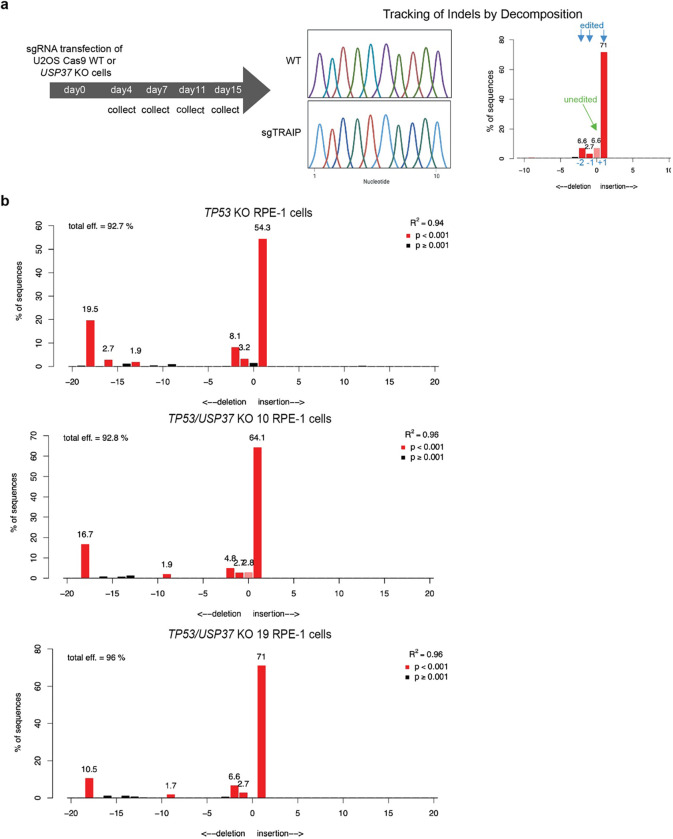
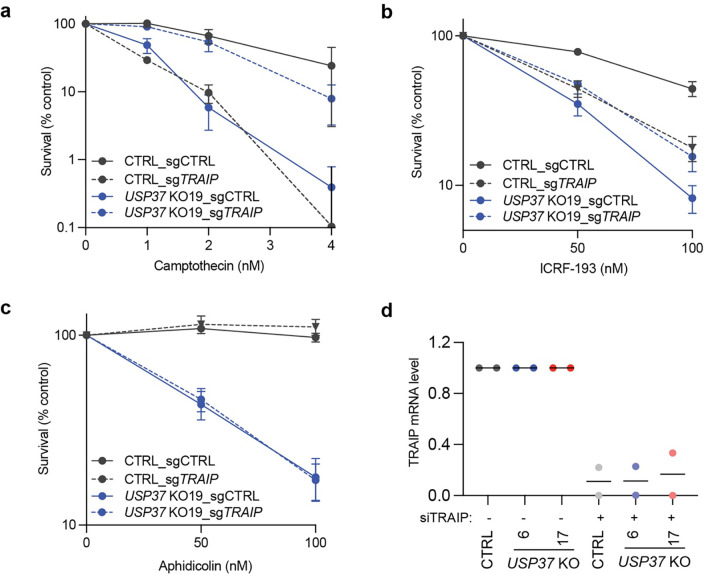
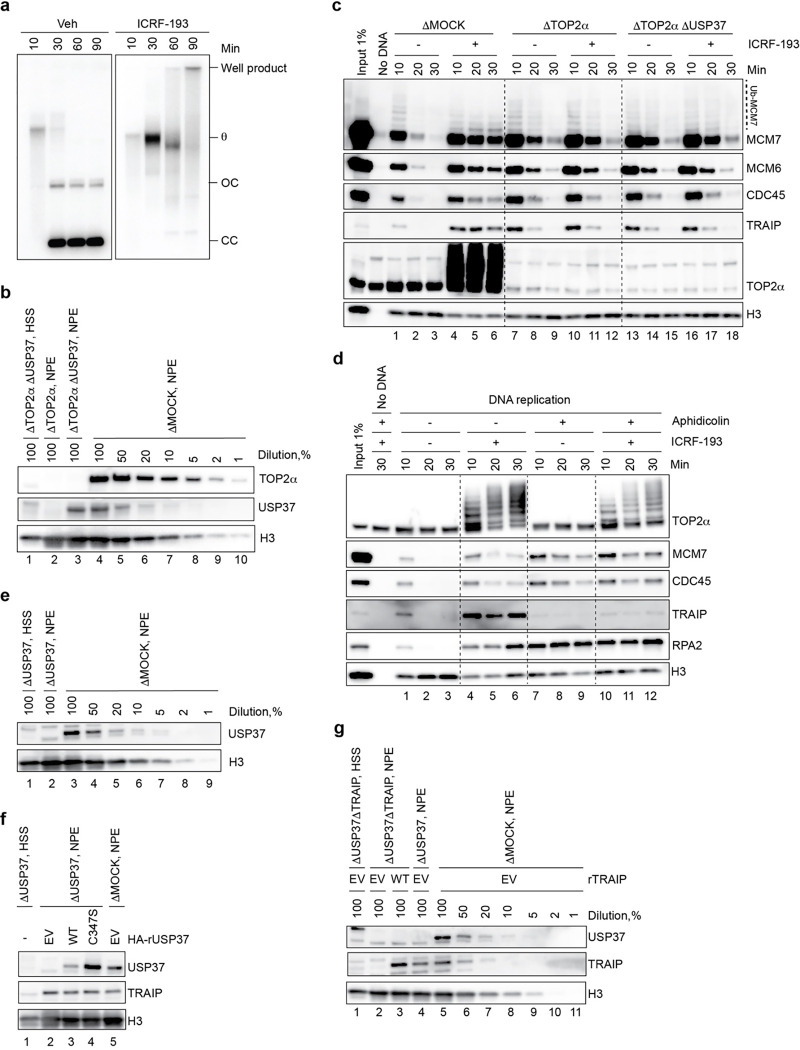
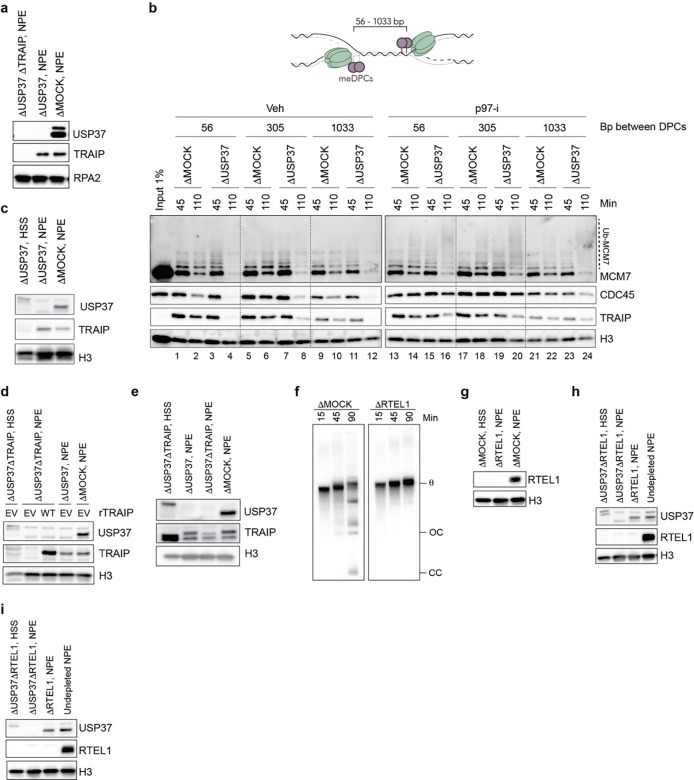
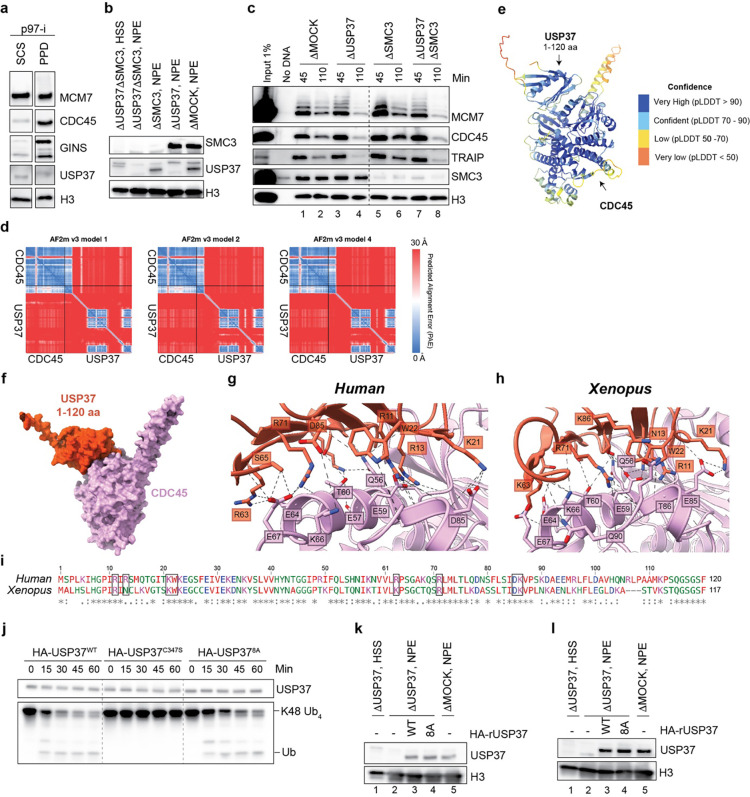
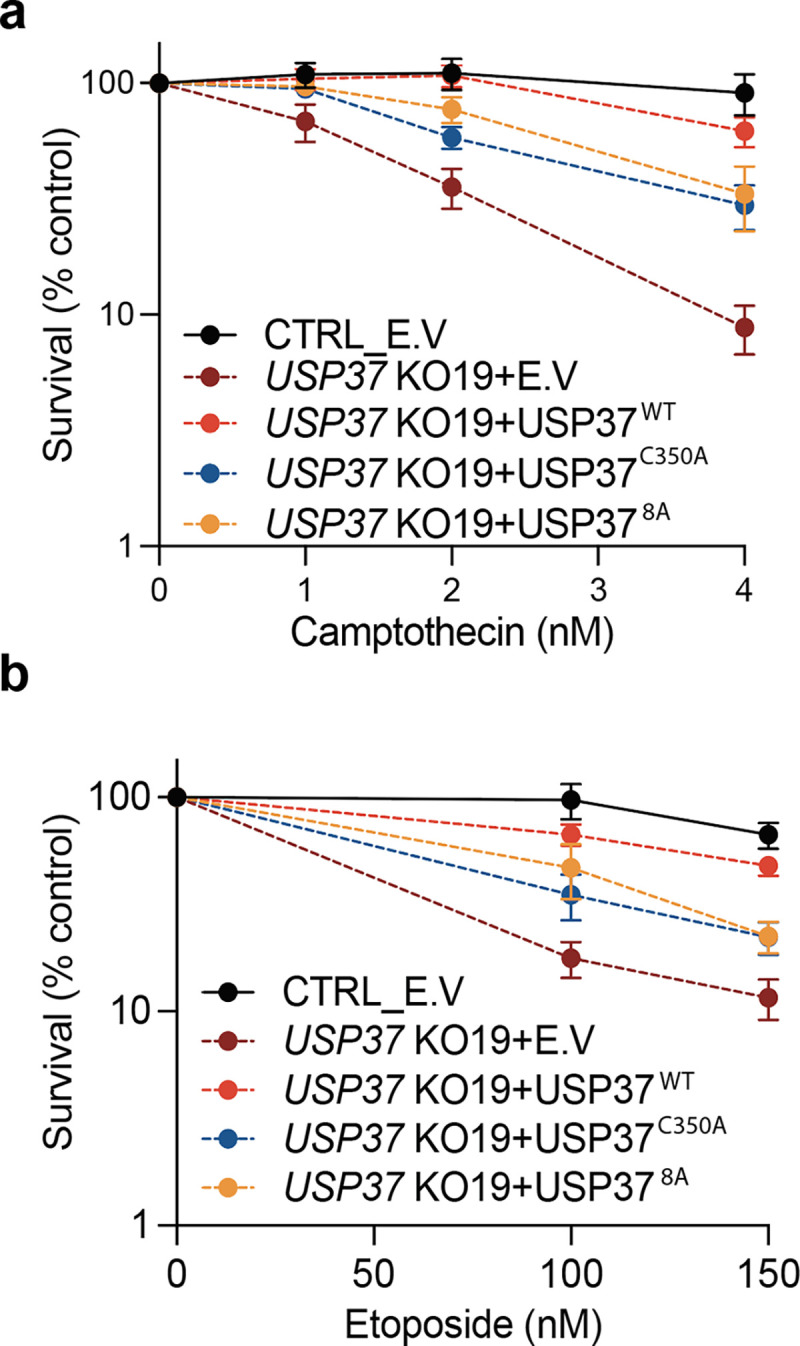
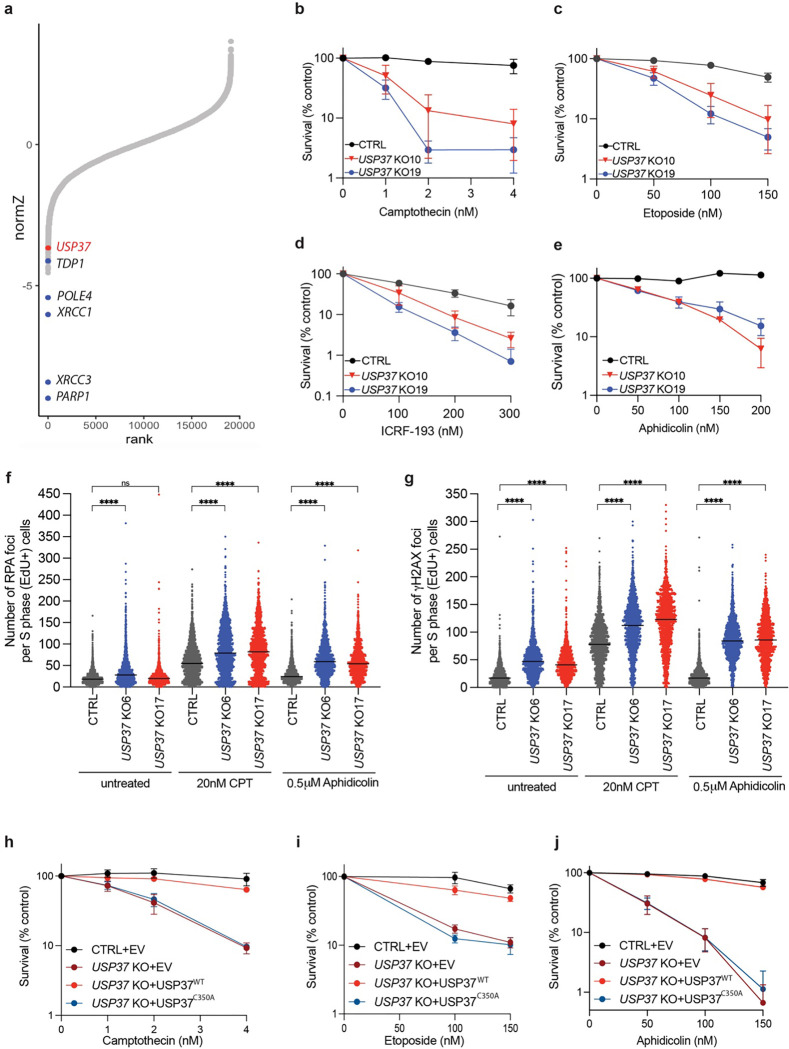
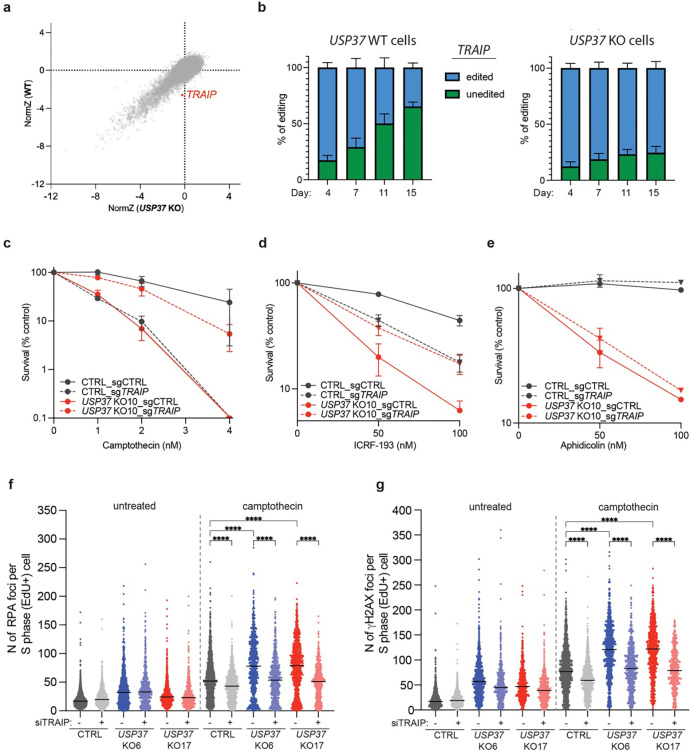
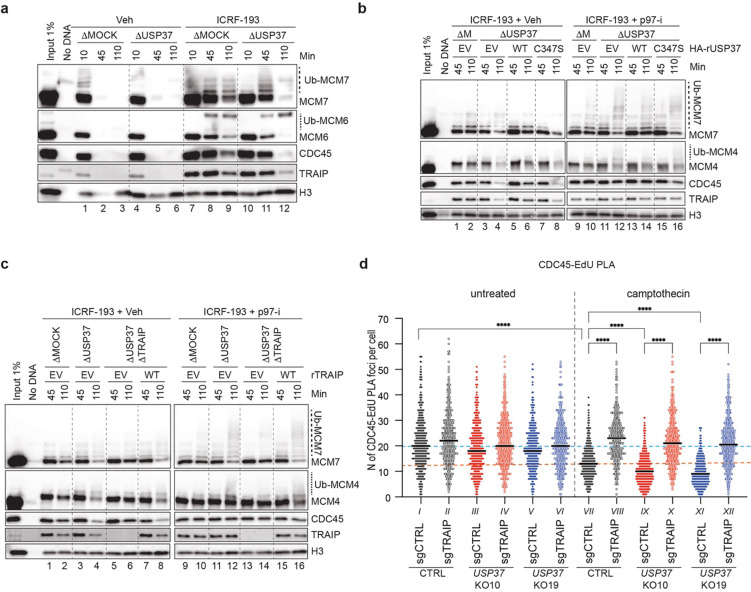
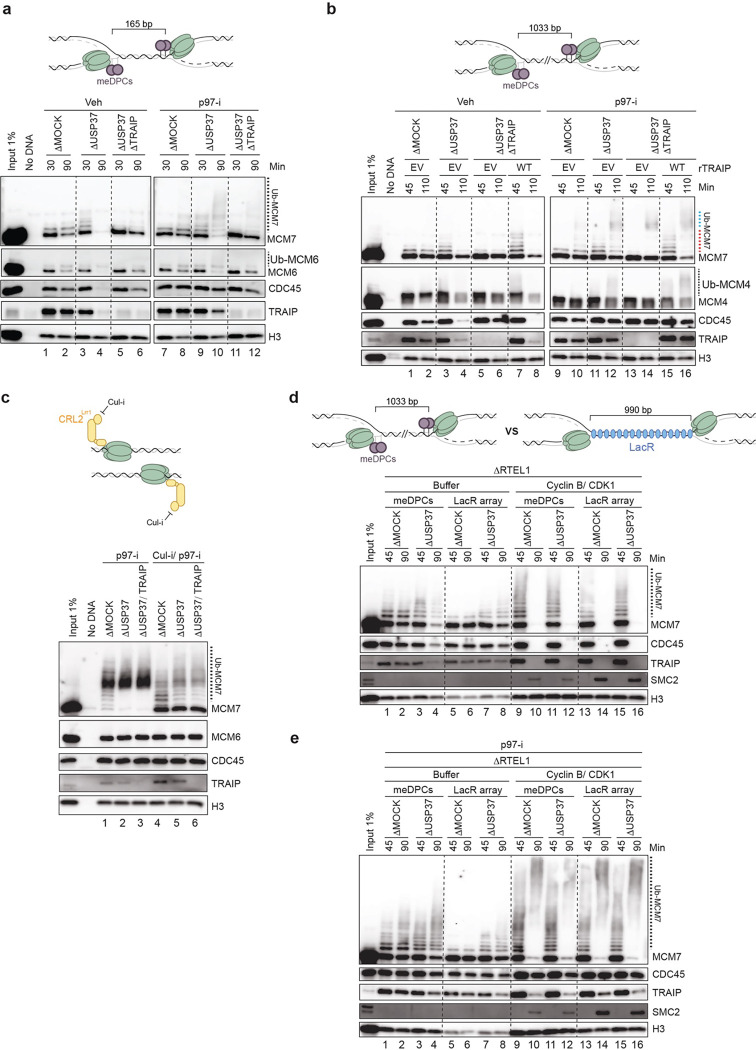
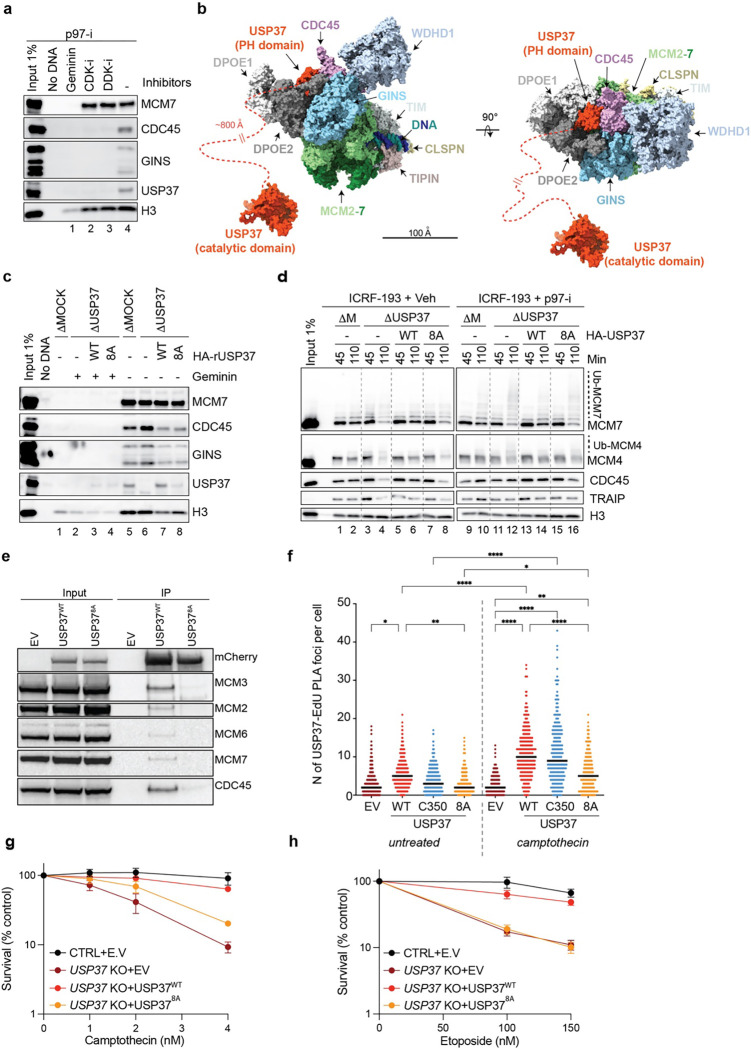
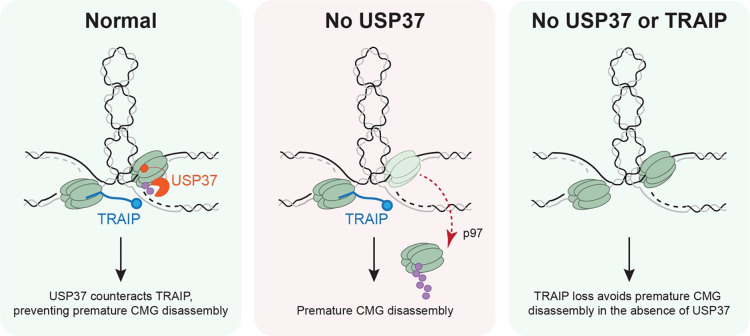
The E3 ubiquitin ligase TRAIP associates with the replisome and helps this molecular machine deal with replication stress. Thus, TRAIP promotes DNA inter-strand crosslink repair by triggering the disassembly of CDC45-MCM2-7-GINS (CMG) helicases that have converged on these lesions. However, disassembly of single CMGs that have stalled temporarily would be deleterious, suggesting that TRAIP must be carefully regulated. Here, we demonstrate that human cells lacking the de-ubiquitylating enzyme USP37 are hypersensitive to topoisomerase poisons and other replication stress-inducing agents. We further show that TRAIP loss rescues the hypersensitivity of USP37 knockout cells to topoisomerase inhibitors. In Xenopus egg extracts depleted of USP37, TRAIP promotes premature CMG ubiquitylation and disassembly when converging replisomes stall. Finally, guided by AlphaFold-Multimer, we discovered that binding to CDC45 mediates USP37's response to topological stress. In conclusion, we propose that USP37 protects genome stability by preventing TRAIP-dependent CMG unloading when replication stress impedes timely termination.
Conflict of interest statement
Competing interests J.C.W. is a co-founder of MOMA Therapeutics, in which he has a financial interest. S.P.J. is Chief Research Officer (part time) at Insmed Innovation UK. Ltd. and founding partner of Ahren Innovation Capital LLP. He is a board member and chair of Scientific Advisory Board of Mission Therapeutics Ltd. and is a consultant and shareholder of Inflex Ltd. The remaining authors declare no competing interests.
Figures
References
-
- Costa A. & Diffley J. F. X. The Initiation of Eukaryotic DNA Replication. Annu Rev Biochem 91, 107–131 (2022). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous