

This is a preprint.
USP37 prevents unscheduled replisome unloading through MCM complex deubiquitination
- PMID: 39282338
- PMCID: PMC11398414
- DOI: 10.1101/2024.09.03.610997
USP37 prevents unscheduled replisome unloading through MCM complex deubiquitination
Update in
-
USP37 prevents unscheduled replisome unloading through MCM complex deubiquitination.Nat Commun. 2025 May 16;16(1):4575. doi: 10.1038/s41467-025-59770-7. Nat Commun. 2025. PMID: 40379725 Free PMC article.
Abstract
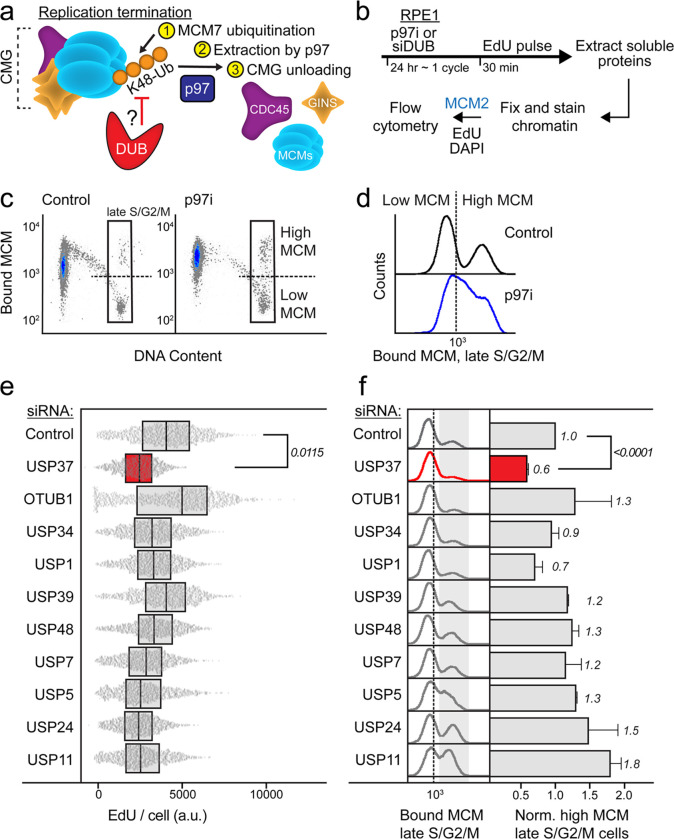
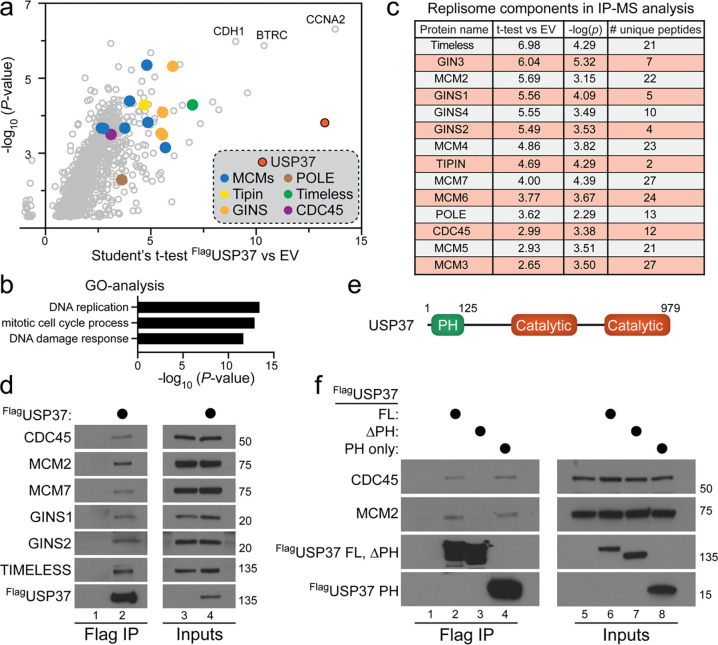
The CMG helicase (CDC45-MCM2-7-GINS) unwinds DNA as a component of eukaryotic replisomes. Replisome (dis)assembly is tightly coordinated with cell cycle progression to ensure genome stability. However, factors that prevent premature CMG unloading and replisome disassembly are poorly described. Since disassembly is catalyzed by ubiquitination, deubiquitinases (DUBs) represent attractive candidates for safeguarding against untimely and deleterious CMG unloading. We combined a targeted loss-of-function screen with quantitative, single-cell analysis to identify human USP37 as a key DUB preventing replisome disassembly. We demonstrate that USP37 maintains active replisomes on S-phase chromatin and promotes normal cell cycle progression. Proteomics and enzyme assays revealed USP37 interacts with the CMG complex to deubiquitinate MCM7, thus antagonizing replisome disassembly. Significantly, USP37 protects normal epithelial cells from oncoprotein-induced replication stress. Our findings reveal USP37 to be critical to the maintenance of replisomes in S-phase and suggest USP37-targeting as a potential strategy for treating malignancies with defective DNA replication control.
Keywords: CMG; DNA replication; genome integrity; replisome; ubiquitin.
Conflict of interest statement
COMPETING INTERESTS The Brown laboratory receives research funding from Amgen. The remaining authors declare no competing interests.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous