

Infection length and host environment influence on Plasmodium falciparum dry season reservoir
- PMID: 39284949
- PMCID: PMC11473648
- DOI: 10.1038/s44321-024-00127-w
Infection length and host environment influence on Plasmodium falciparum dry season reservoir
Abstract
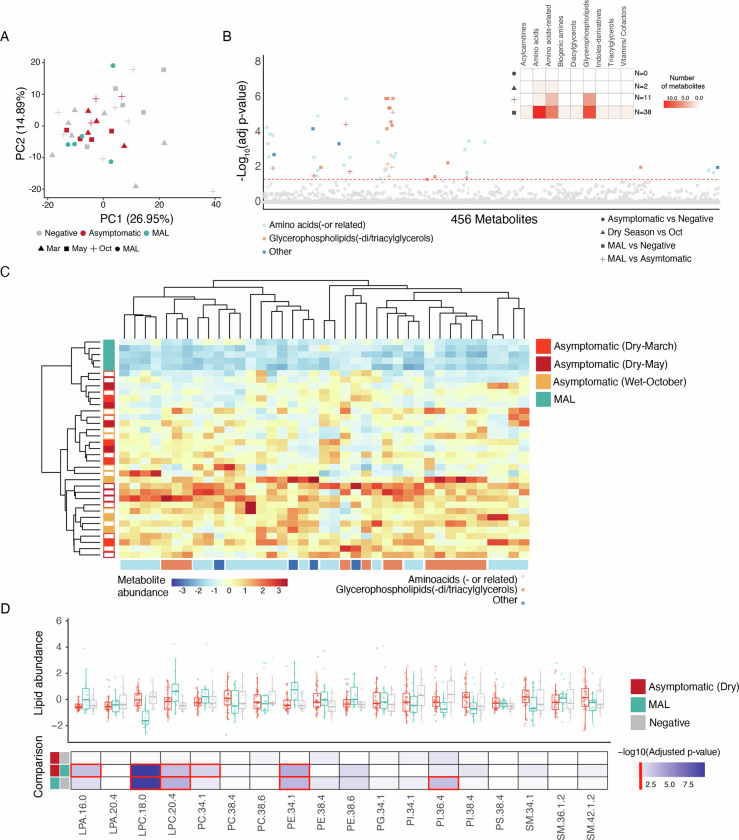
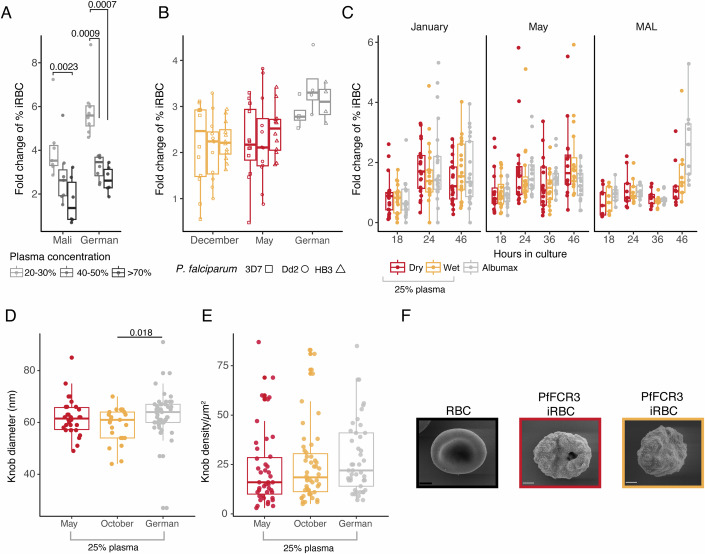
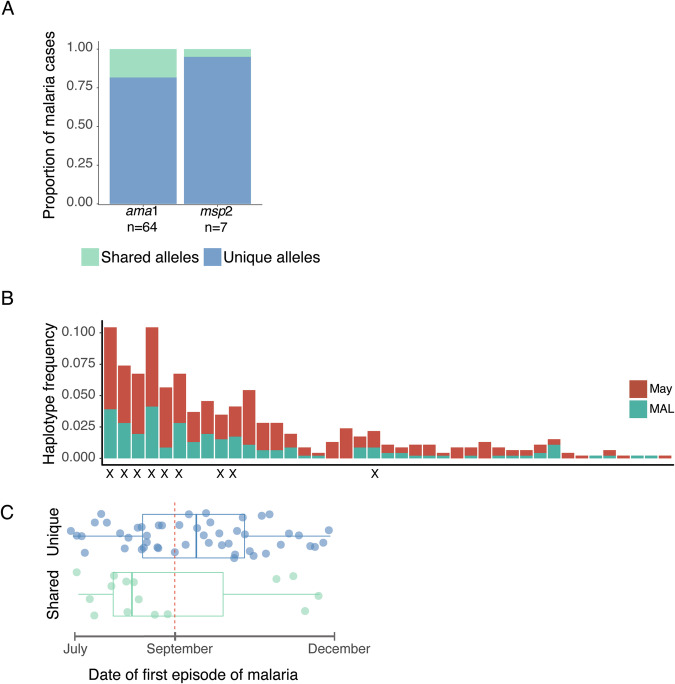
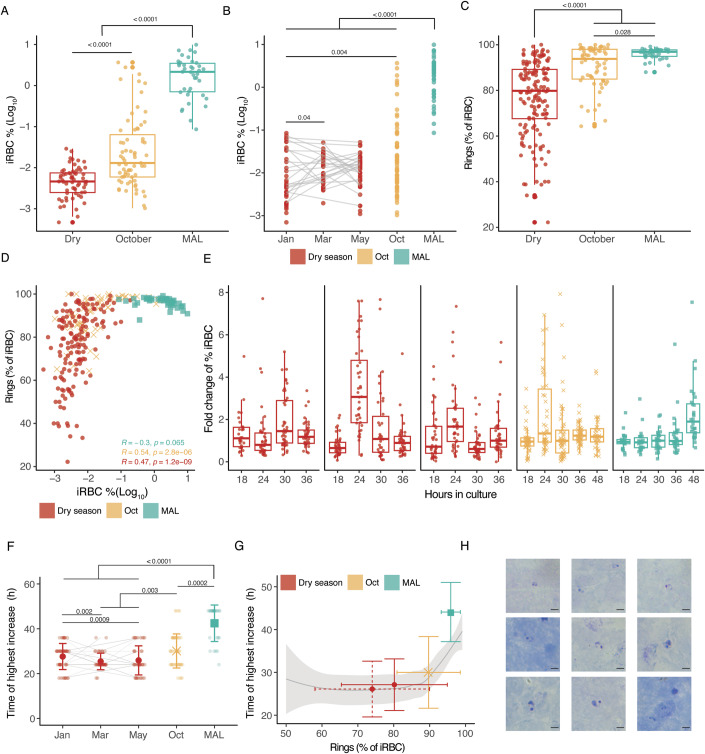
Persistence of malaria parasites in asymptomatic hosts is crucial in areas of seasonally-interrupted transmission, where P. falciparum bridges wet seasons months apart. During the dry season, infected erythrocytes exhibit extended circulation with reduced cytoadherence, increasing the risk of splenic clearance of infected cells and hindering parasitaemia increase. However, what determines parasite persistence for long periods of time remains unknown. Here, we investigated whether seasonality affects plasma composition so that P. falciparum can detect and adjust to changing serological cues; or if alternatively, parasite infection length dictates clinical presentation and persistency. Data from Malian children exposed to alternating ~6-month wet and dry seasons show that plasma composition is unrelated to time of year in non-infected children, and that carrying P. falciparum only minimally affects plasma constitution in asymptomatic hosts. Parasites persisting in the blood of asymptomatic children from the dry into the ensuing wet season rarely if ever appeared to cause malaria in their hosts as seasons changed. In vitro culture in the presence of plasma collected in the dry or the wet seasons did not affect parasite development, replication or host-cell remodelling. The absence of a parasite-encoded sensing mechanism was further supported by the observation of similar features in P. falciparum persisting asymptomatically in the dry season and parasites in age- and sex-matched asymptomatic children in the wet season. Conversely, we show that P. falciparum clones transmitted early in the wet season had lower chance of surviving until the end of the following dry season, contrasting with a higher likelihood of survival of clones transmitted towards the end of the wet season, allowing for the re-initiation of transmission. We propose that the decreased virulence observed in persisting parasites during the dry season is not due to the parasites sensing ability, nor is it linked to a decreased capacity for parasite replication but rather a consequence decreased cytoadhesion associated with infection length.
Keywords: Asymptomatic; Dry Season; Infection Length; Malaria; Sensing.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Andolina C, Ramjith J, Rek J, Lanke K, Okoth J, Grignard L, Arinaitwe E, Briggs J, Bailey J, Aydemir O et al (2023) Plasmodium falciparum gametocyte carriage in longitudinally monitored incident infections is associated with duration of infection and human host factors. Sci Rep. 13:7072 - PMC - PubMed
-
- Andrade CM, Fleckenstein H, Thomson-Luque R, Doumbo S, Lima NF, Anderson C, Hibbert J, Hopp CS, Tran TM, Li S et al (2020) Increased circulation time of Plasmodium falciparum underlies persistent asymptomatic infection in the dry season. Nat Med 26:1929–1940 - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
