

Targeting SNRNP200-induced splicing dysregulation offers an immunotherapy opportunity for glycolytic triple-negative breast cancer
- PMID: 39285160
- PMCID: PMC11405407
- DOI: 10.1038/s41421-024-00715-7
Targeting SNRNP200-induced splicing dysregulation offers an immunotherapy opportunity for glycolytic triple-negative breast cancer
Abstract
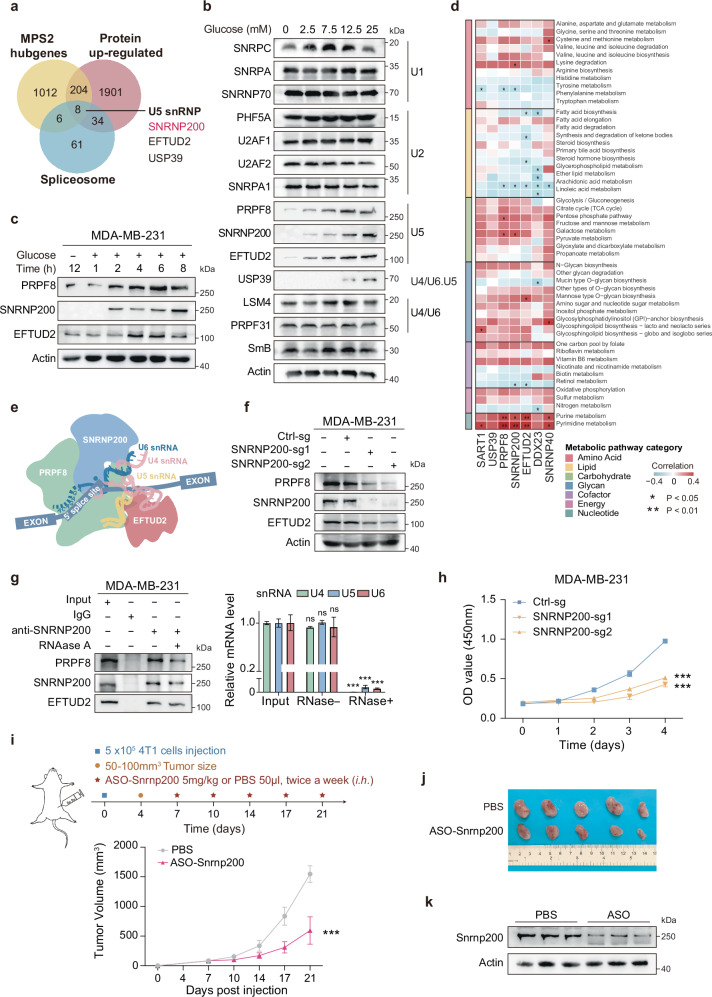
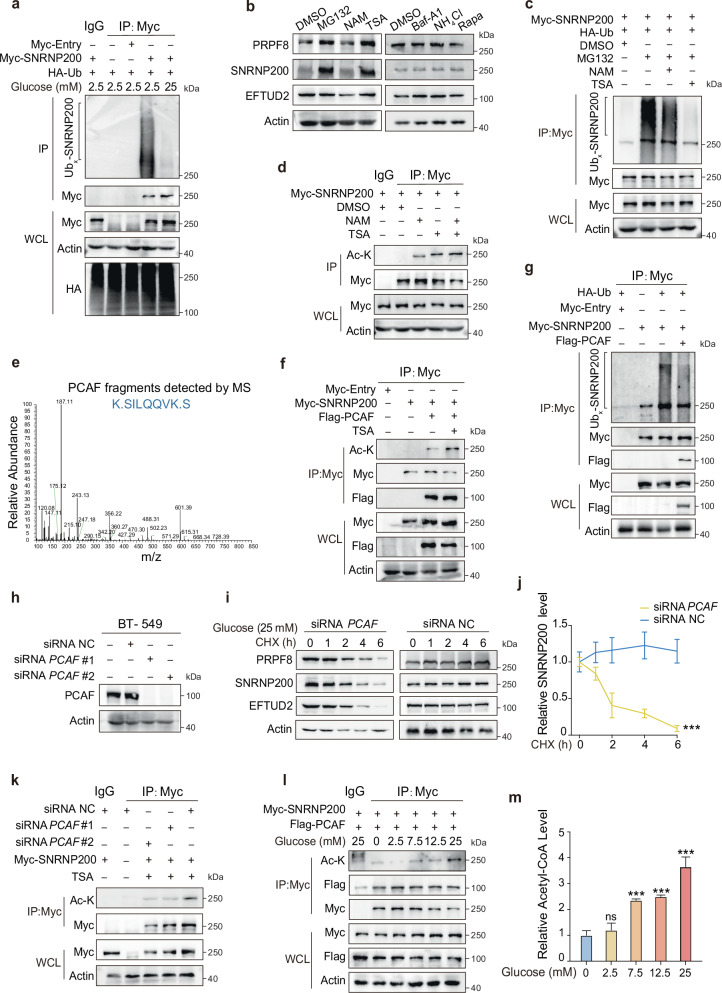
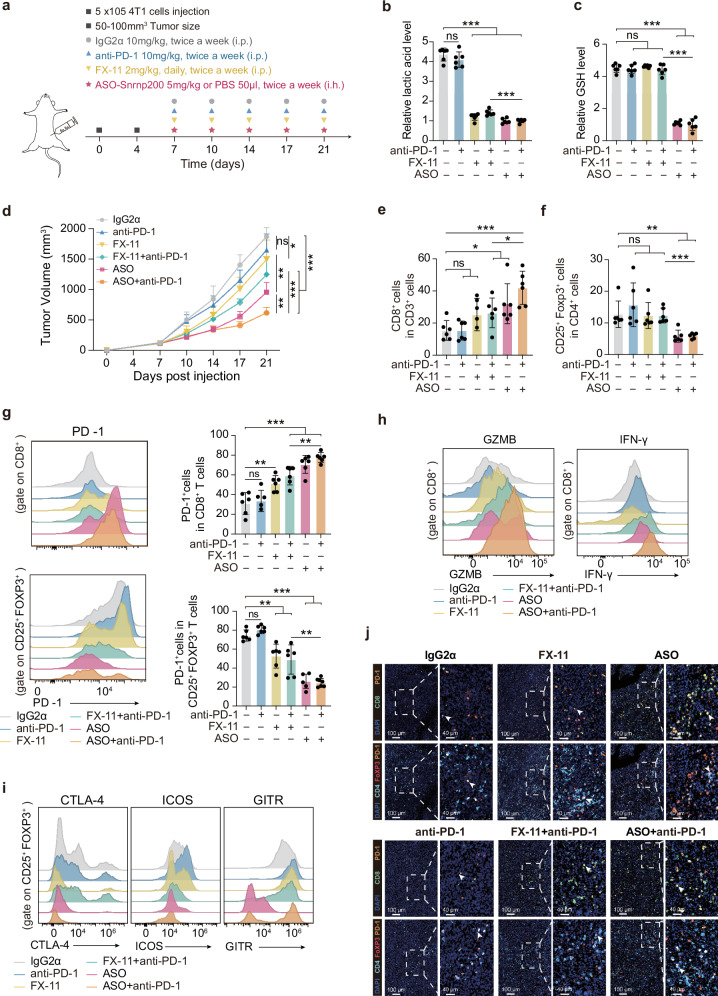
Metabolic dysregulation is prominent in triple-negative breast cancer (TNBC), yet therapeutic strategies targeting cancer metabolism are limited. Here, utilizing multiomics data from our TNBC cohort (n = 465), we demonstrated widespread splicing deregulation and increased spliceosome abundance in the glycolytic TNBC subtype. We identified SNRNP200 as a crucial mediator of glucose-driven metabolic reprogramming. Mechanistically, glucose induces acetylation at SNRNP200 K1610, preventing its proteasomal degradation. Augmented SNRNP200 then facilitates splicing key metabolic enzyme-encoding genes (GAPDH, ALDOA, and GSS), leading to increased lactic acid and glutathione production. Targeting SNRNP200 with antisense oligonucleotide therapy impedes tumor metabolism and enhances the efficacy of anti-PD-1 therapy by activating intratumoral CD8+ T cells while suppressing regulatory T cells. Clinically, higher SNRNP200 levels indicate an inferior response to immunotherapy in glycolytic TNBCs. Overall, our study revealed the intricate interplay between RNA splicing and metabolic dysregulation, suggesting an innovative combination strategy for immunotherapy in glycolytic TNBCs.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Grants and funding
- 81872137/National Natural Science Foundation of China (National Science Foundation of China)
- 82072917/National Natural Science Foundation of China (National Science Foundation of China)
- 82272957/National Natural Science Foundation of China (National Science Foundation of China)
- 82303735/National Natural Science Foundation of China (National Science Foundation of China)
- 22DZ2204400/Shanghai Science and Technology Development Foundation (Shanghai Science and Technology Development Fund)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
