

Atomic vacancies of molybdenum disulfide nanoparticles stimulate mitochondrial biogenesis
- PMID: 39289340
- PMCID: PMC11408498
- DOI: 10.1038/s41467-024-52276-8
Atomic vacancies of molybdenum disulfide nanoparticles stimulate mitochondrial biogenesis
Abstract
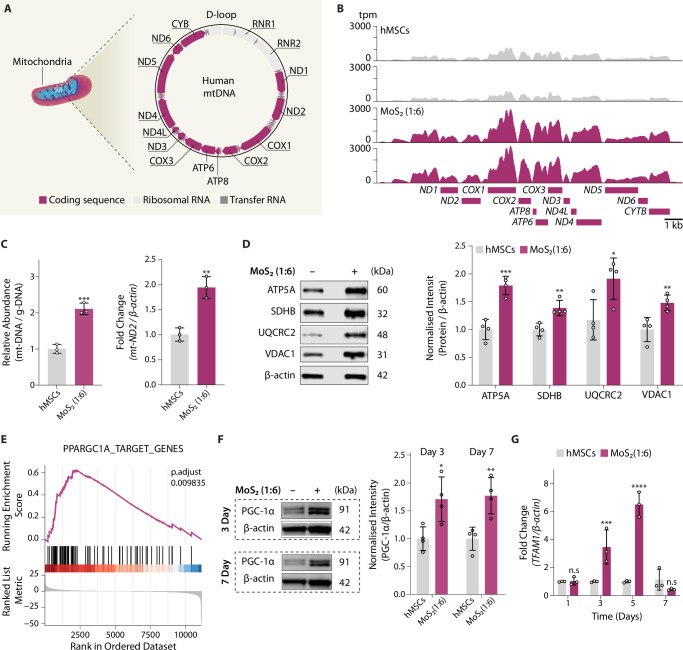
Diminished mitochondrial function underlies many rare inborn errors of energy metabolism and contributes to more common age-associated metabolic and neurodegenerative disorders. Thus, boosting mitochondrial biogenesis has been proposed as a potential therapeutic approach for these diseases; however, currently we have a limited arsenal of compounds that can stimulate mitochondrial function. In this study, we designed molybdenum disulfide (MoS2) nanoflowers with predefined atomic vacancies that are fabricated by self-assembly of individual two-dimensional MoS2 nanosheets. Treatment of mammalian cells with MoS2 nanoflowers increased mitochondrial biogenesis by induction of PGC-1α and TFAM, which resulted in increased mitochondrial DNA copy number, enhanced expression of nuclear and mitochondrial-DNA encoded genes, and increased levels of mitochondrial respiratory chain proteins. Consistent with increased mitochondrial biogenesis, treatment with MoS2 nanoflowers enhanced mitochondrial respiratory capacity and adenosine triphosphate production in multiple mammalian cell types. Taken together, this study reveals that predefined atomic vacancies in MoS2 nanoflowers stimulate mitochondrial function by upregulating the expression of genes required for mitochondrial biogenesis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare the following competing interests. A.K.G., K.A.S., V.M.G., and I.S. have filed a patent application related to the work presented in this article. The remaining authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- R35 GM152102/GM/NIGMS NIH HHS/United States
- R01GM143630/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R21 NS121945/NS/NINDS NIH HHS/United States
- R01 GM143630/GM/NIGMS NIH HHS/United States
- DE032031/U.S. Department of Health & Human Services | NIH | National Institute of Dental and Craniofacial Research (NIDCR)
- R01 DE032031/DE/NIDCR NIH HHS/United States
- R35GM152102/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- RP230204/Cancer Prevention and Research Institute of Texas (Cancer Prevention Research Institute of Texas)
- NS121945/U.S. Department of Health & Human Services | NIH | National Institute of Neurological Disorders and Stroke (NINDS)
- W81XWH2210932/United States Department of Defense | United States Army | Army Medical Command | Congressionally Directed Medical Research Programs (CDMRP)
- EB026265/U.S. Department of Health & Human Services | NIH | National Institute of Biomedical Imaging and Bioengineering (NIBIB)
- R01 CA282251/CA/NCI NIH HHS/United States
- CA282251/U.S. Department of Health & Human Services | NIH | NCI | Division of Cancer Epidemiology and Genetics, National Cancer Institute (National Cancer Institute Division of Cancer Epidemiology and Genetics)
- DP2 EB026265/EB/NIBIB NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
