

Molecular mechanisms of thalidomide effectiveness on COVID-19 patients explained: ACE2 is a new ΔNp63α target gene
- PMID: 39294414
- PMCID: PMC11525293
- DOI: 10.1007/s00109-024-02485-x
Molecular mechanisms of thalidomide effectiveness on COVID-19 patients explained: ACE2 is a new ΔNp63α target gene
Abstract
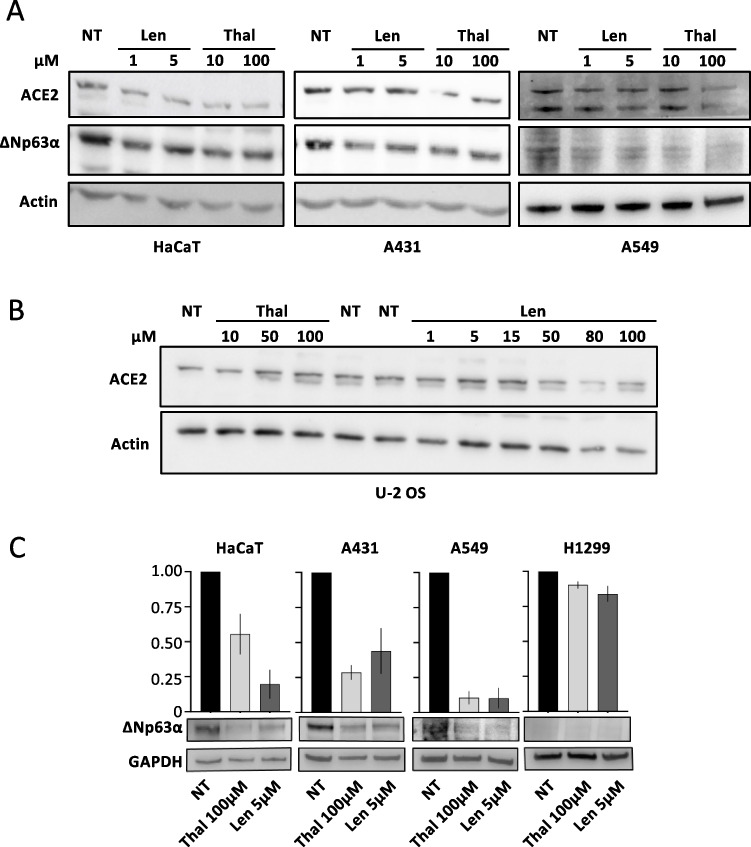
COVID-19 pandemic is caused by the SARS-CoV-2 virus, whose internalization and infection are mediated by the angiotensin-converting enzyme 2 (ACE2). The identification of novel approaches to tackle this step is instrumental for the development of therapies for the management of COVID-19 and other diseases with a similar mechanism of infection. Thalidomide, a drug sadly known for its teratogenic effects, has potent immunomodulatory and anti-inflammatory properties. Treatment with this drug has been shown to improve the immune functions of COVID-19 patients and proposed for the management of COVID-19 in clinical practice through drug repositioning. Here, we investigated the molecular details linking thalidomide to ACE2 and COVID-19, showing that in conditions mimicking SARS-CoV-2-associated cytokine storm, the transcription factor ΔNp63α and ACE2 are stabilized, and IL-8 production is increased. In such conditions, we found p63 to bind to and regulate the expression of the ACE2 gene. We previously showed that ΔNp63α is degraded upon thalidomide treatment and now found that treatment with this drug-or with its analogue lenalidomide-downregulates ACE2 in a p63-dependent manner. Finally, we found that thalidomide treatment reduces in vitro infection by pseudo-SARS-CoV-2, a baculovirus pseudotyped with the SARS-CoV-2 spike protein. Overall, we propose the dual effect of thalidomide in reducing SARS-CoV-2 viral re-entry and inflammation through p63 degradation to weaken SARS-CoV-2 entry into host cells and mitigate lung inflammation, making it a valuable option in clinical management of COVID-19. KEY MESSAGES: Thalidomide treatment results in p63-dependent ACE2 downregulation. ACE2 is a p63 transcriptional target. Thalidomide reduces the "cytokine storm" associated to COVID-19. Thalidomide prevents viral re-entry of SARS-CoV-2 by p63-dependent ACE2 downregulation. Thalidomide is a modulator of SARS-CoV-2 or other ACE2-dependent infections. ACE2 is modulated by a pharmacological substance.
Keywords: ACE2; COVID 19; SARS-CoV-2; Thalidomide; ΔNp63α.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, Schiergens TS, Herrler G, Wu NH, Nitsche A et al (2020) SARS-CoV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell 181(2):271–280. 10.1016/j.cell.2020.02.052 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
