

Long-lived lung megakaryocytes contribute to platelet recovery in thrombocytopenia models
- PMID: 39302653
- PMCID: PMC11563682
- DOI: 10.1172/JCI181111
Long-lived lung megakaryocytes contribute to platelet recovery in thrombocytopenia models
Abstract
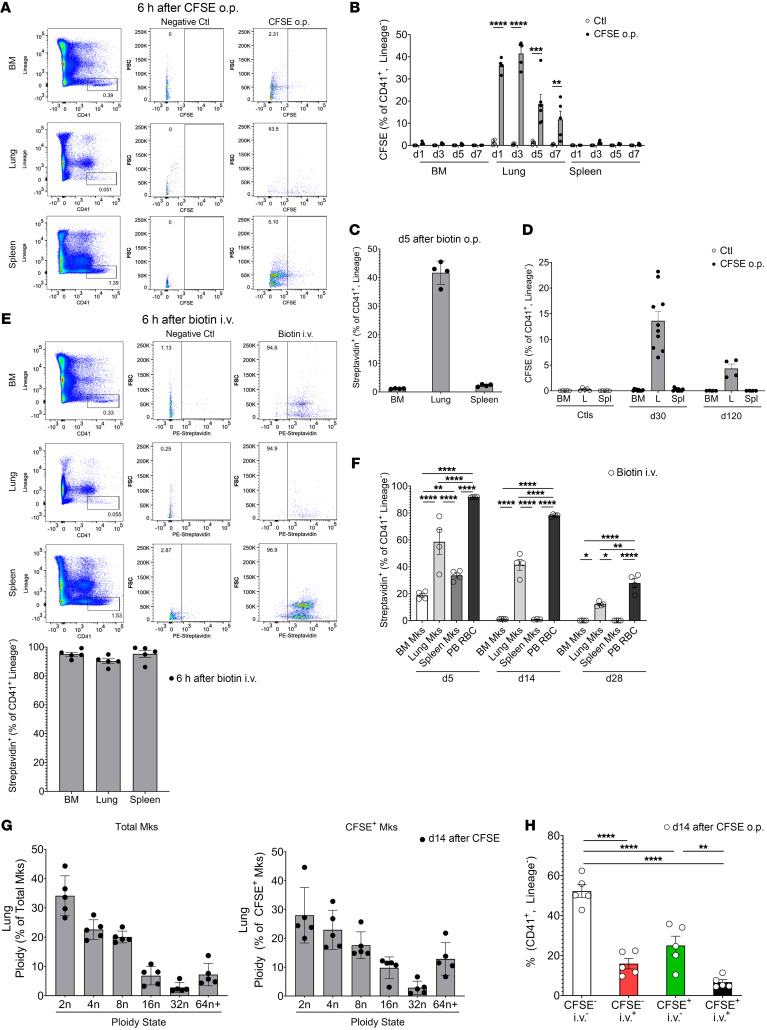
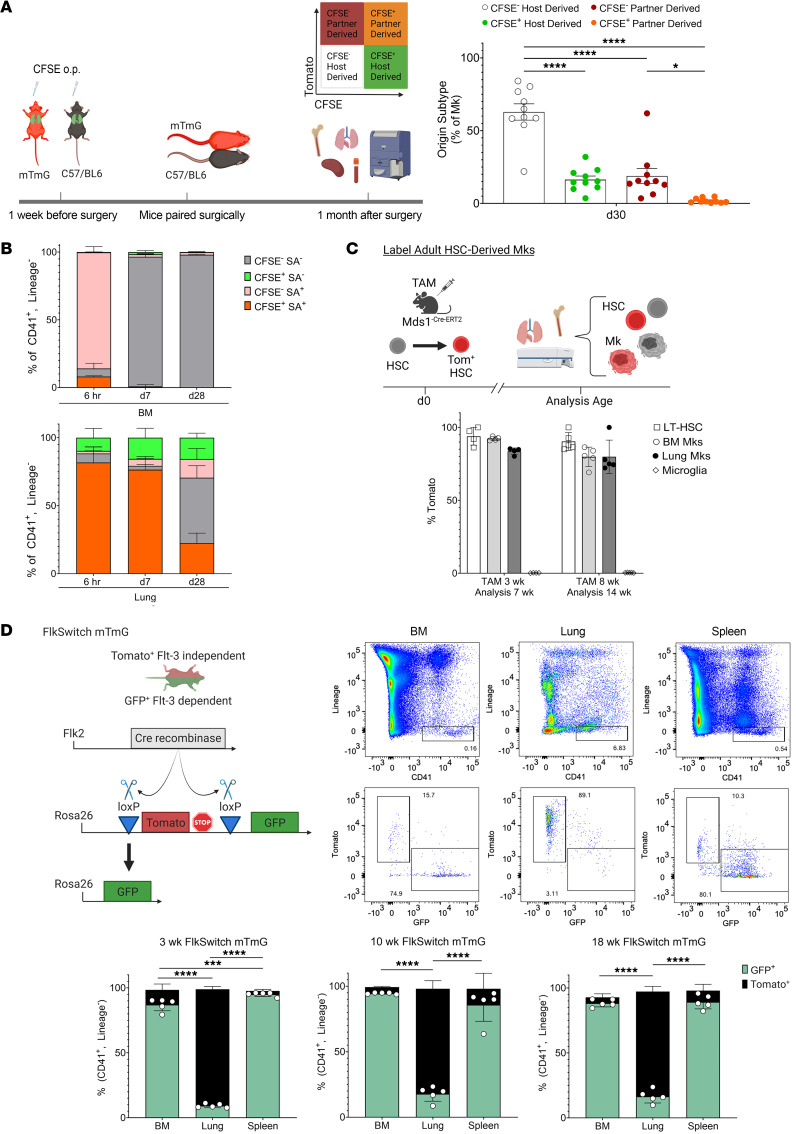
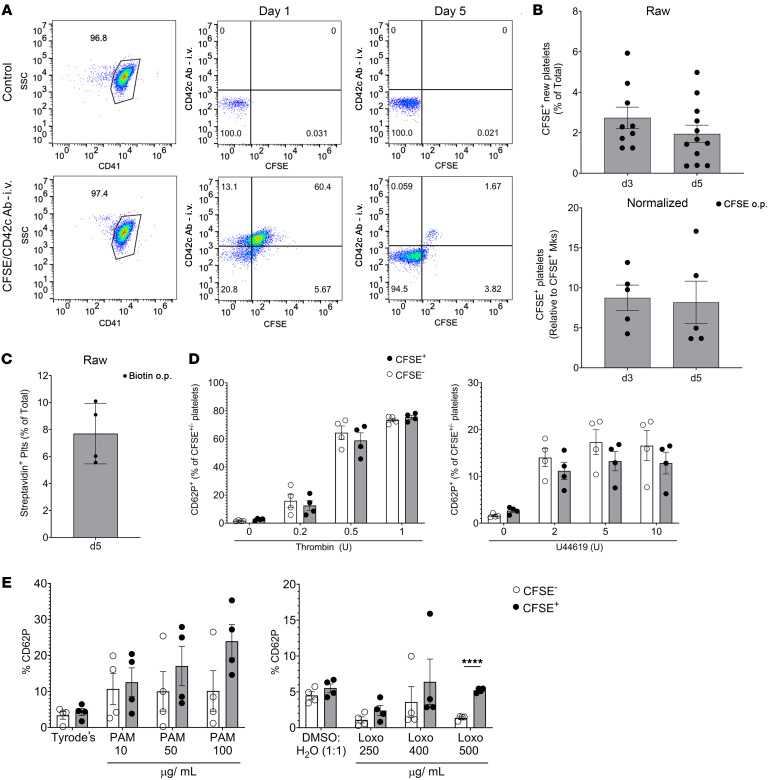
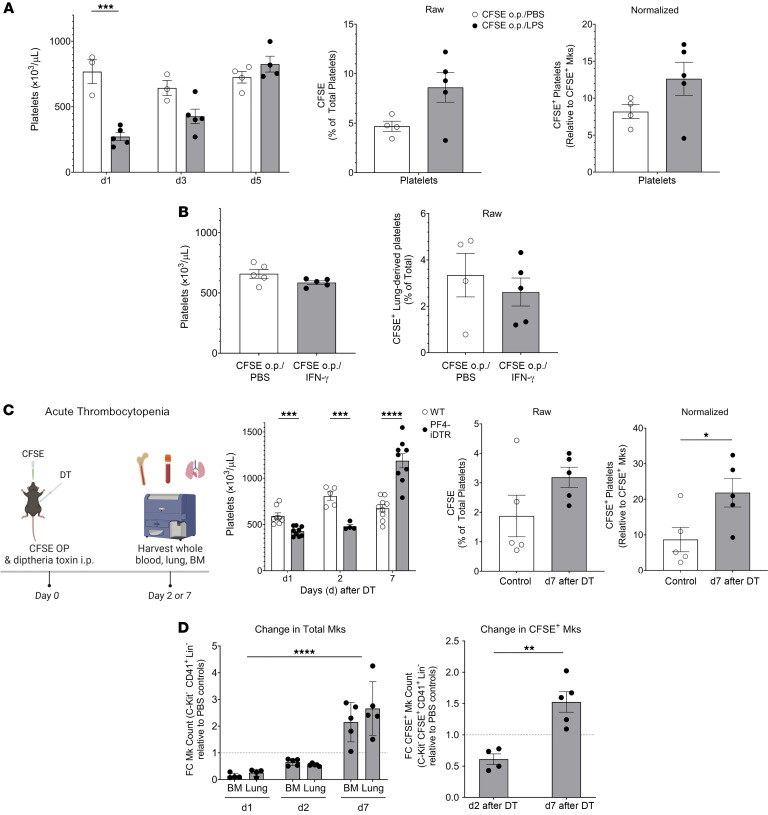
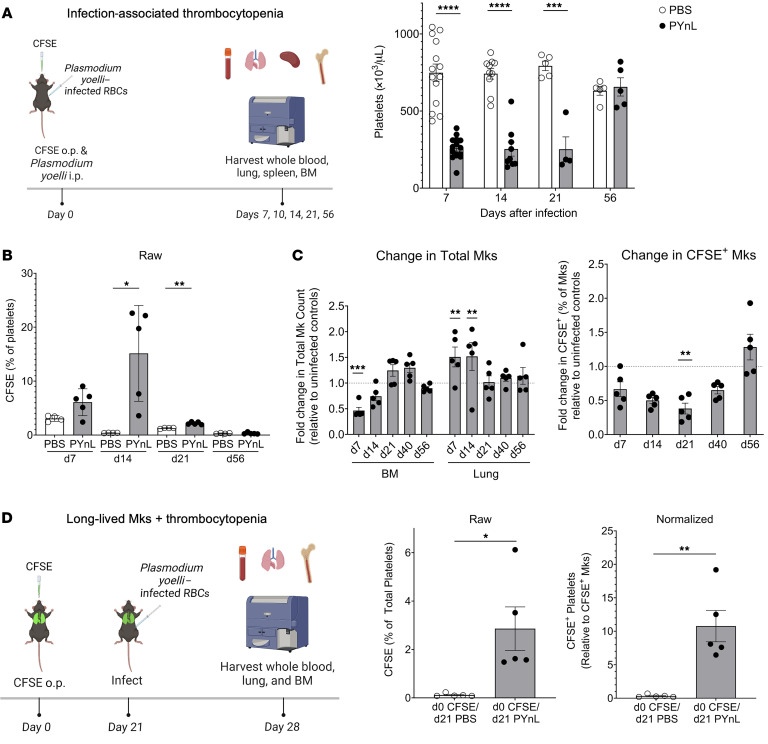
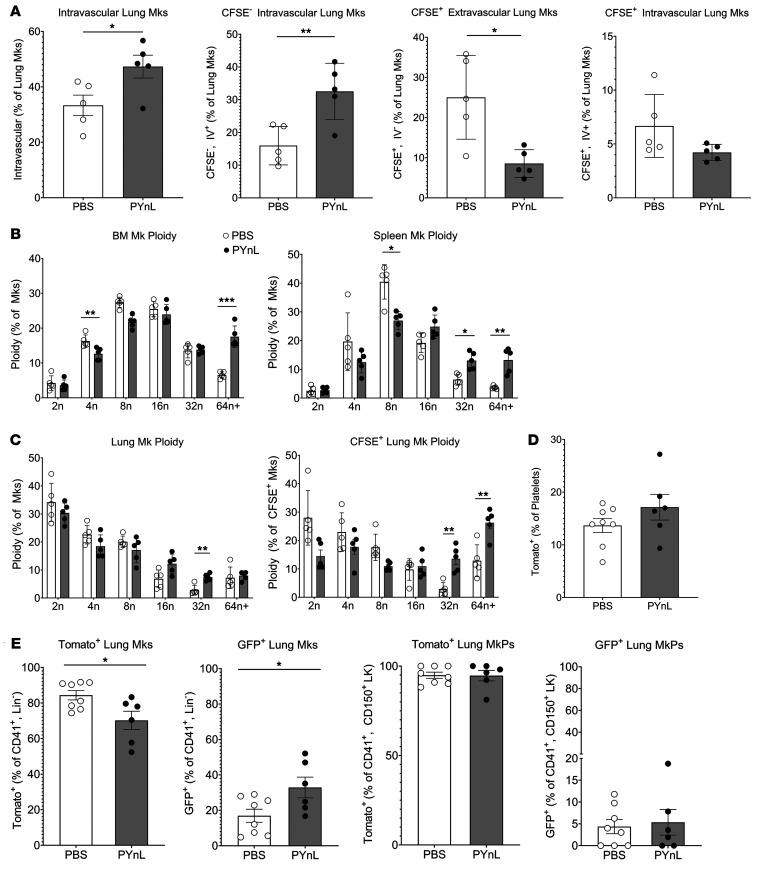
Lung megakaryocytes (Mks) are largely extravascular with an immune phenotype (1). Because bone marrow (BM) Mks are short lived, it has been assumed that extravascular lung Mks are constantly "seeded" from the BM. To investigate lung Mk origins and how origin affects their functions, we developed methods to specifically label lung Mks using CFSE dye and biotin delivered via the oropharyngeal route. Labeled lung Mks were present for up to 4 months, while BM Mks had a lifespan of less than 1 week. In a parabiosis model, lung Mks were partially replaced over 1 month from a circulating source. Unlike tissue-resident macrophages, using MDS1-Cre-ERT2 TdTomato mice, we found that lung Mks arose from hematopoietic stem cells. However, studies with FlkSwitch mTmG mice showed that lung Mks were derived from a Flt3-independent lineage that did not go through a multipotent progenitor. CFSE labeling to track lung Mk-derived platelets showed that approximately 10% of circulating platelets were derived from lung-resident Mks at steady state, but in sterile thrombocytopenia this was doubled (~20%). Lung-derived platelets were similarly increased in a malaria infection model (Plasmodium yoelii) typified by thrombocytopenia. These studies indicate that lung Mks arise from a Flt3- BM source, are long-lived, and contribute more platelets during thrombocytopenia.
Keywords: Hematology; Platelets.
Conflict of interest statement
Figures
Comment in
- The lung is a megakaryocyte outpost that can defend against thrombocytopenic attack
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
