

Two-dose priming immunization amplifies humoral immunity by synchronizing vaccine delivery with the germinal center response
- PMID: 39303017
- PMCID: PMC11492009
- DOI: 10.1126/sciimmunol.adl3755
Two-dose priming immunization amplifies humoral immunity by synchronizing vaccine delivery with the germinal center response
Abstract
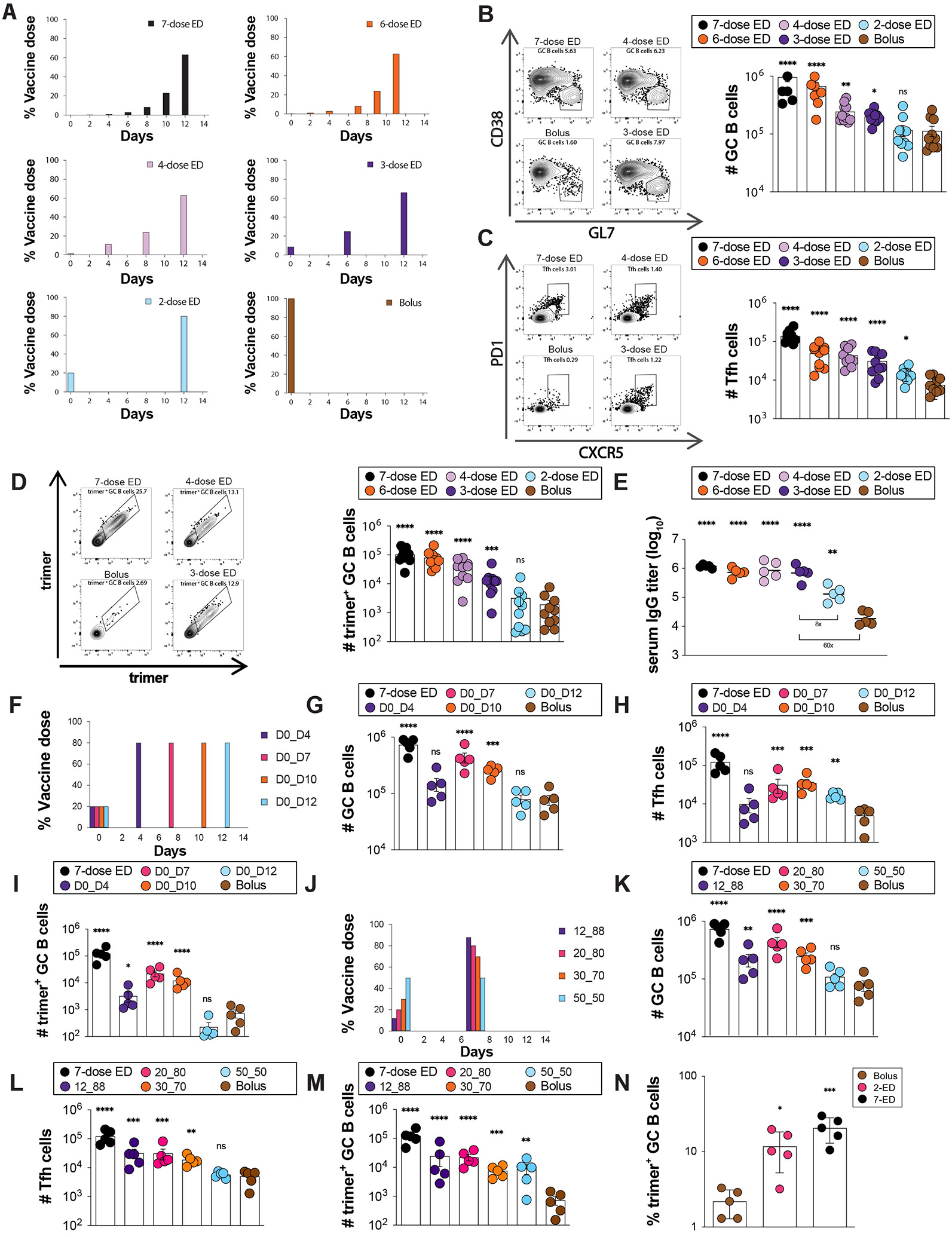
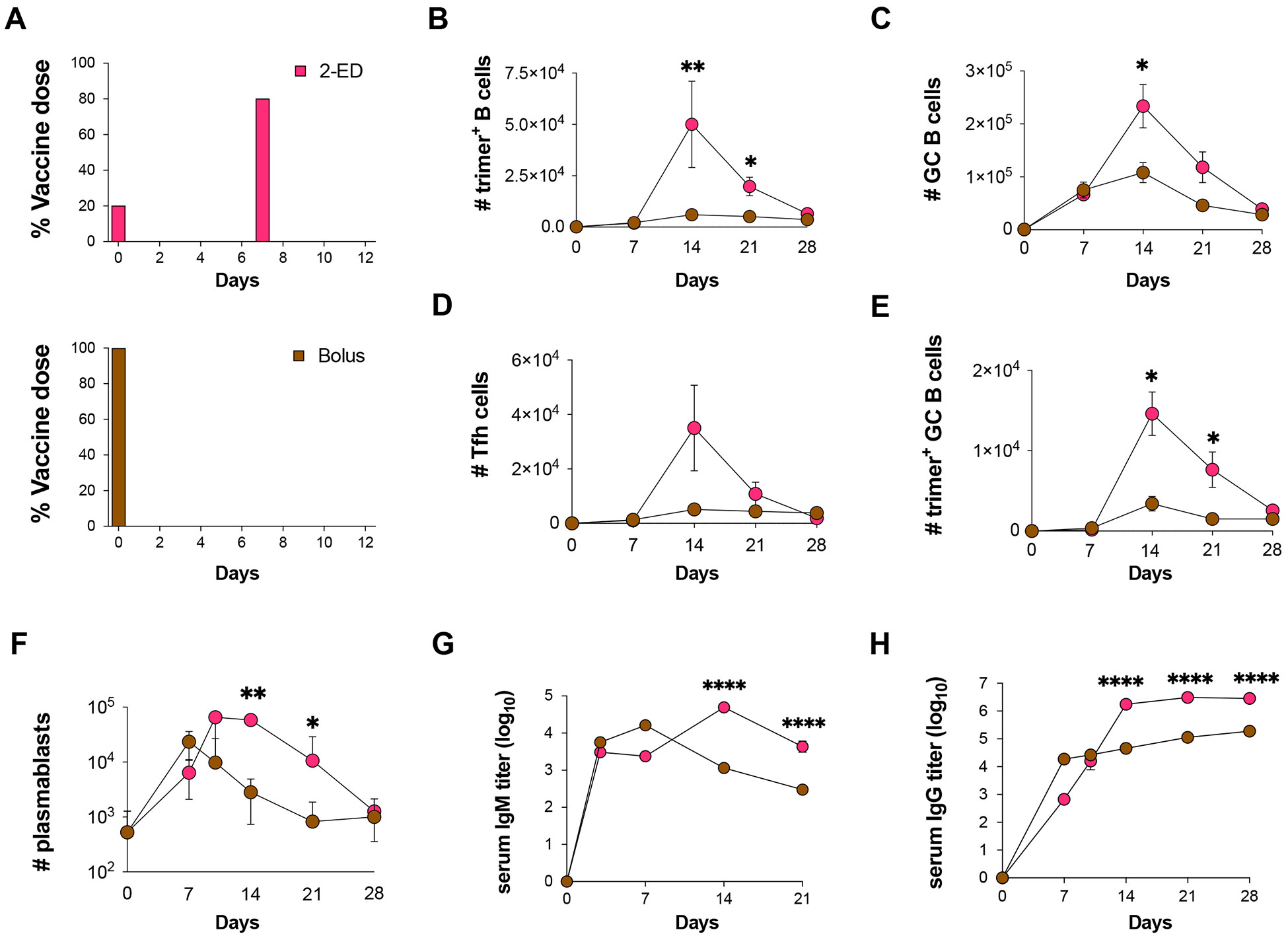
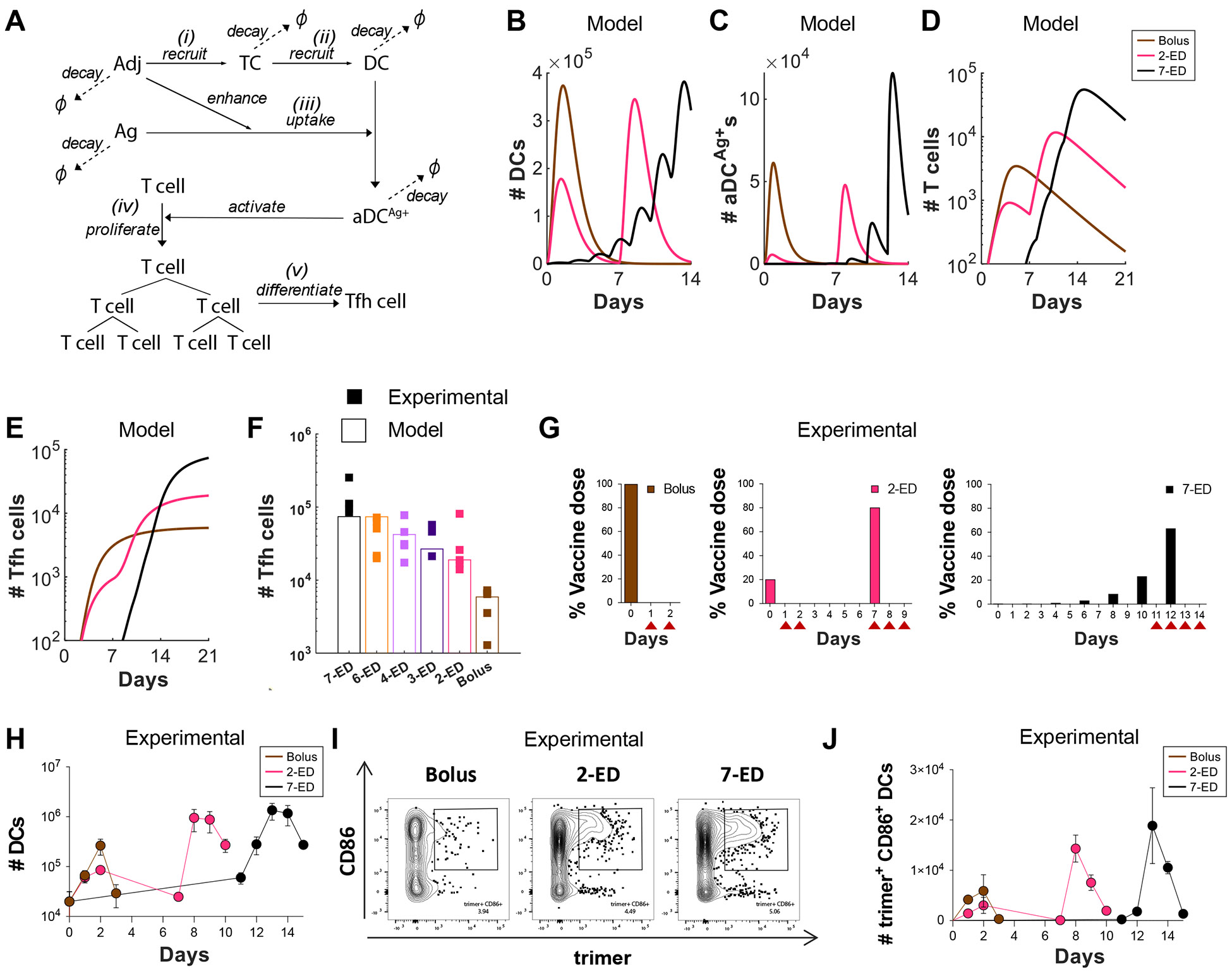
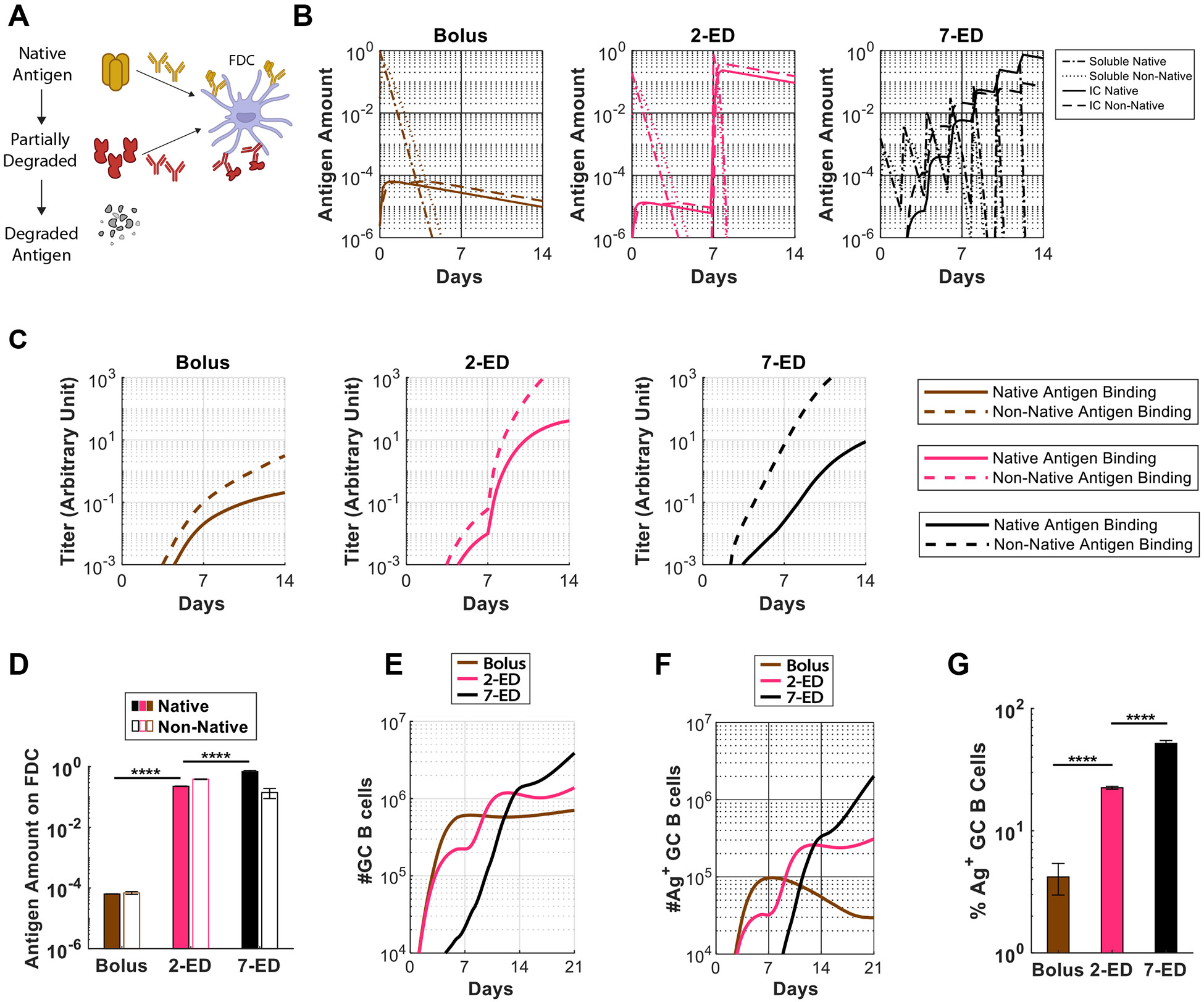
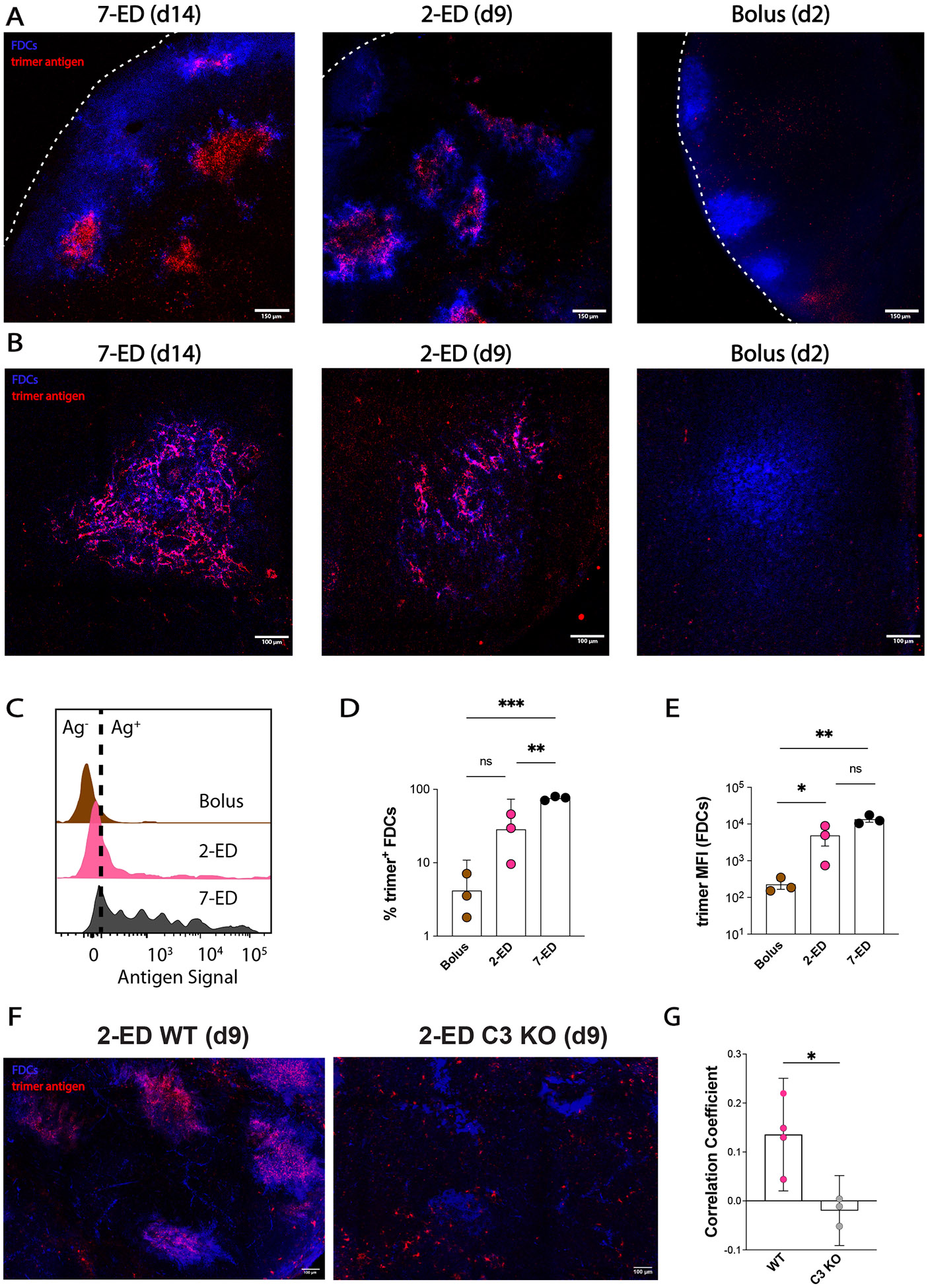
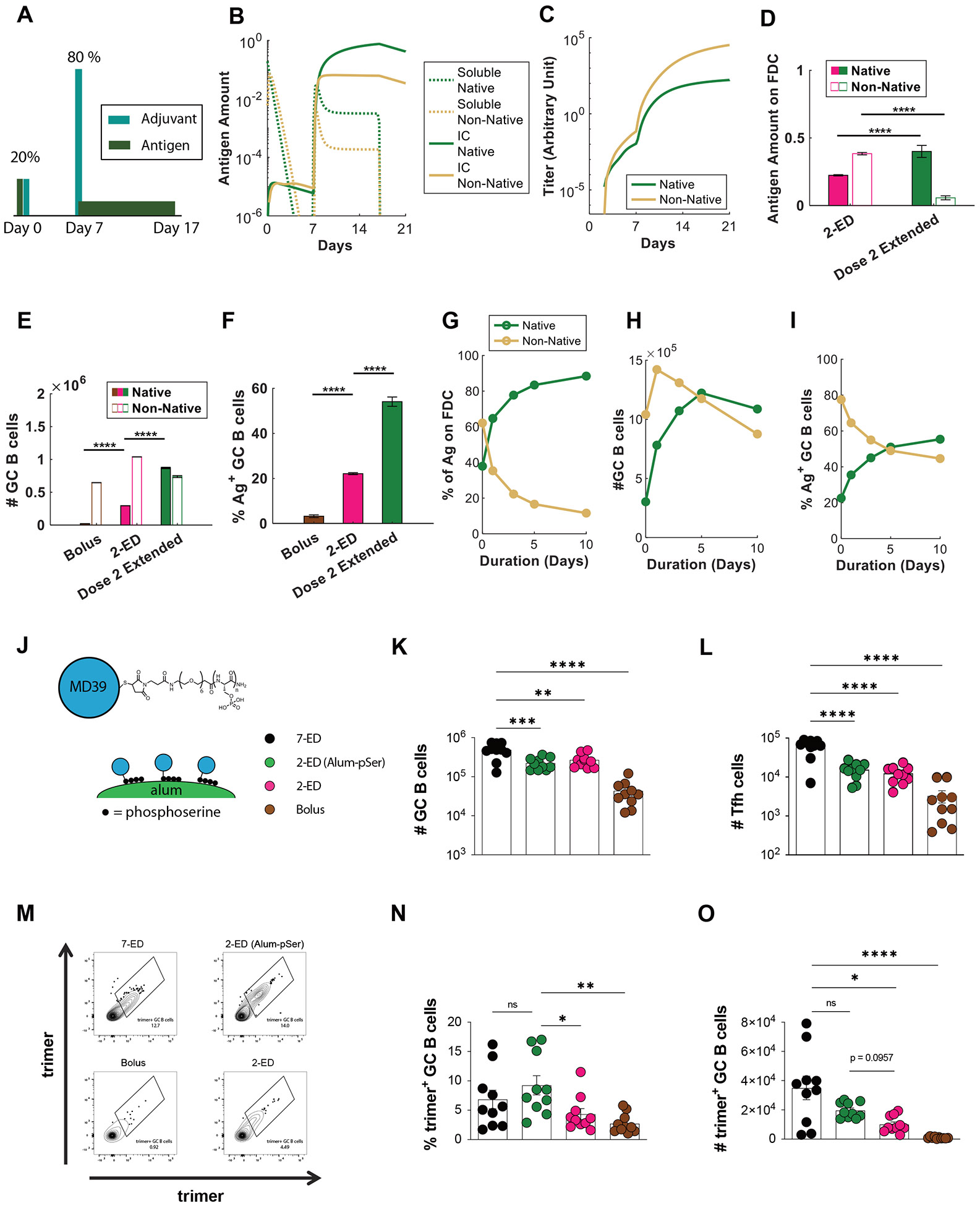
Prolonging exposure to subunit vaccines during the primary immune response enhances humoral immunity. Escalating-dose immunization (EDI), administering vaccines every other day in an increasing pattern over 2 weeks, is particularly effective but challenging to implement clinically. Here, using an HIV Env trimer/saponin adjuvant vaccine, we explored simplified EDI regimens and found that a two-shot regimen administering 20% of the vaccine followed by the remaining 80% of the dose 7 days later increased TFH responses 6-fold, antigen-specific germinal center (GC) B cells 10-fold, and serum antibody titers 10-fold compared with bolus immunization. Computational modeling of TFH priming and the GC response suggested that enhanced activation/antigen loading on dendritic cells and increased capture of antigen delivered in the second dose by follicular dendritic cells contribute to these effects, predictions we verified experimentally. These results suggest that a two-shot priming approach can be used to substantially enhance responses to subunit vaccines.
Figures
Update of
-
Two-dose "extended priming" immunization amplifies humoral immune responses by synchronizing vaccine delivery with the germinal center response.bioRxiv [Preprint]. 2023 Nov 21:2023.11.20.563479. doi: 10.1101/2023.11.20.563479. bioRxiv. 2023. Update in: Sci Immunol. 2024 Sep 20;9(99):eadl3755. doi: 10.1126/sciimmunol.adl3755. PMID: 38045401 Free PMC article. Updated. Preprint.
References
-
- Feinberg MB, Uhambo — Twists and Turns on the Journey to an Efficacious HIV-1 Vaccine. New Engl J Med 384, 1157–1159 (2021). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
