

Protein phosphatase PP2Cα S-glutathionylation regulates cell migration
- PMID: 39303918
- PMCID: PMC11530597
- DOI: 10.1016/j.jbc.2024.107784
Protein phosphatase PP2Cα S-glutathionylation regulates cell migration
Abstract
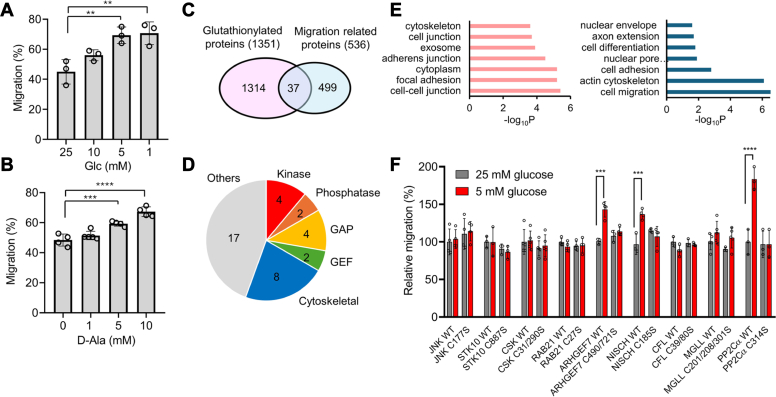
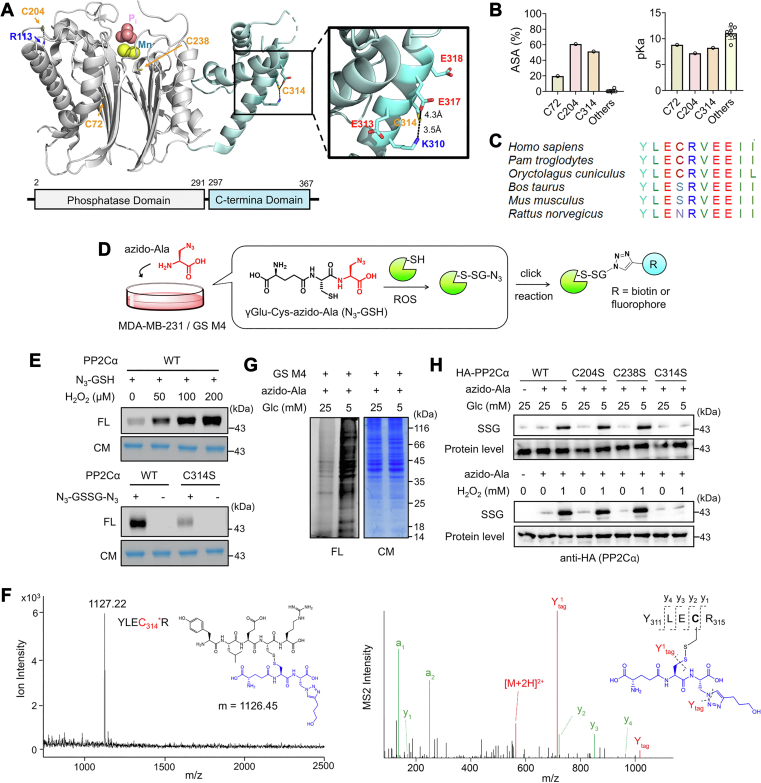
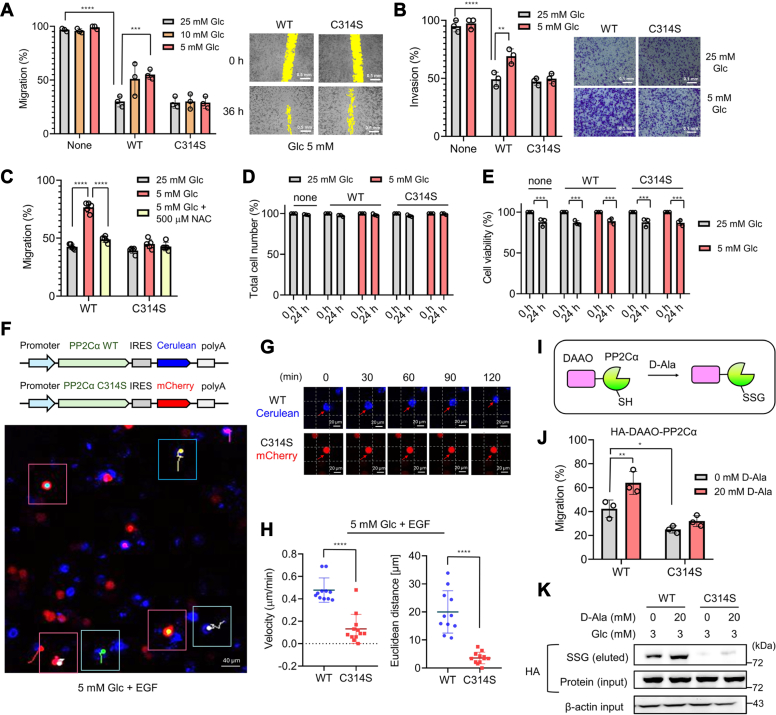
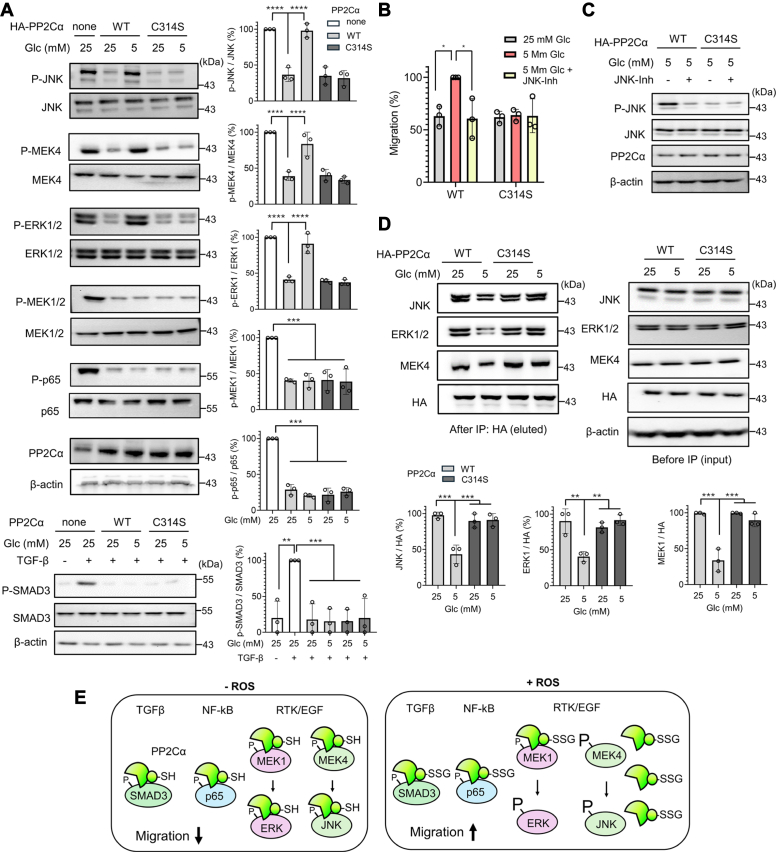
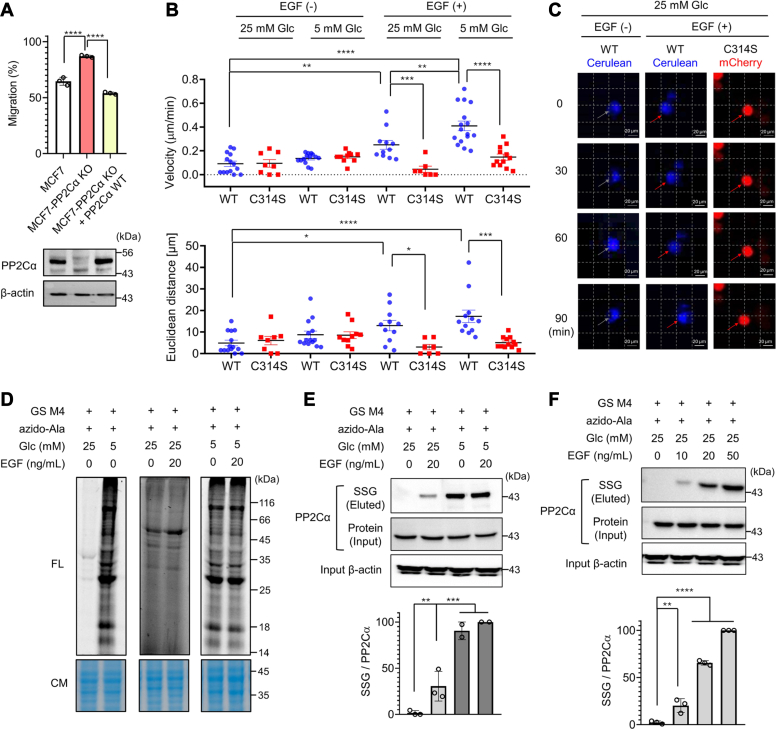
Redox signaling is a fundamental mechanism that controls all major biological processes partly via protein cysteine oxidations, including S-glutathionylation. Despite over 2000 cysteines identified to form S-glutathionylation in databases, the identification of redox cysteines functionally linked to a biological process of interest remains challenging. Here, we demonstrate a strategy combining glutathionylation proteomic database, bioinformatics, and biological screening, which resulted in the identification of S-glutathionylated proteins, including PP2Cα, as redox players of cell migration. We showed that PP2Cα, a prototypical magnesium-dependent serine/threonine phosphatase, is susceptible to S-glutathionylation selectively at nonconserved C314. PP2Cα glutathionylation causes increased migration and invasion of breast cancer cell lines in oxidative stress or upon hydrogen peroxide production. Mechanistically, PP2Cα glutathionylation modulates its protein-protein interactions, activating c-Jun N-terminal kinase and extracellular signal-regulated kinase pathways to elevate migration and invasion. In addition, PP2Cα glutathionylation occurs in response to epidermal growth factor, supporting a serine/threonine phosphatase PP2Cα as a new redox player in growth factor signal transduction.
Keywords: PP2Cα phosphatase; cell migration; epidermal growth factor; glutathionylation; reactive oxygen species; redox signaling.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interests The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Luster A.D., Alon R., von Andrian U.H. Immune cell migration in inflammation: present and future therapeutic targets. Nat. Immunol. 2005;6:1182–1190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
