

HDAC5 controls a hypothalamic STAT5b-TH axis, the sympathetic activation of ATP-consuming futile cycles and adult-onset obesity in male mice
- PMID: 39304061
- PMCID: PMC11481749
- DOI: 10.1016/j.molmet.2024.102033
HDAC5 controls a hypothalamic STAT5b-TH axis, the sympathetic activation of ATP-consuming futile cycles and adult-onset obesity in male mice
Abstract
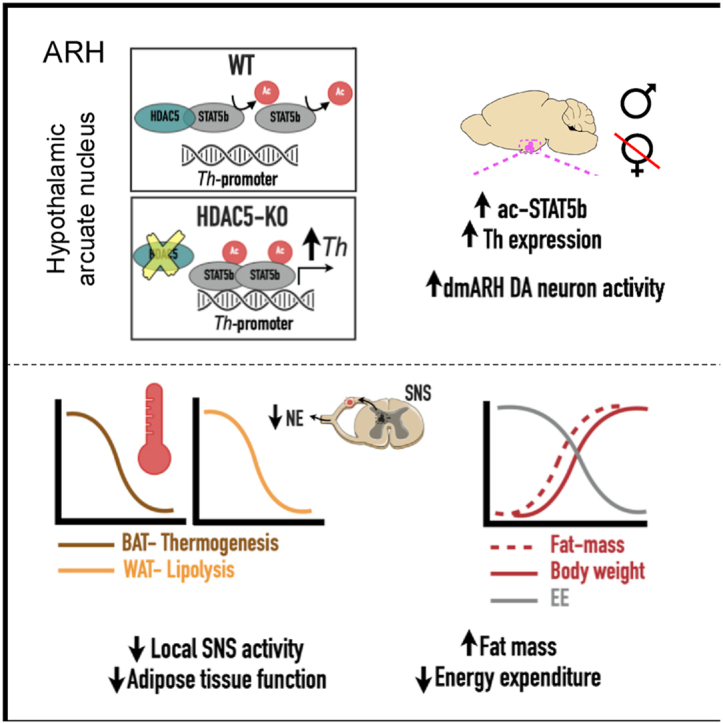
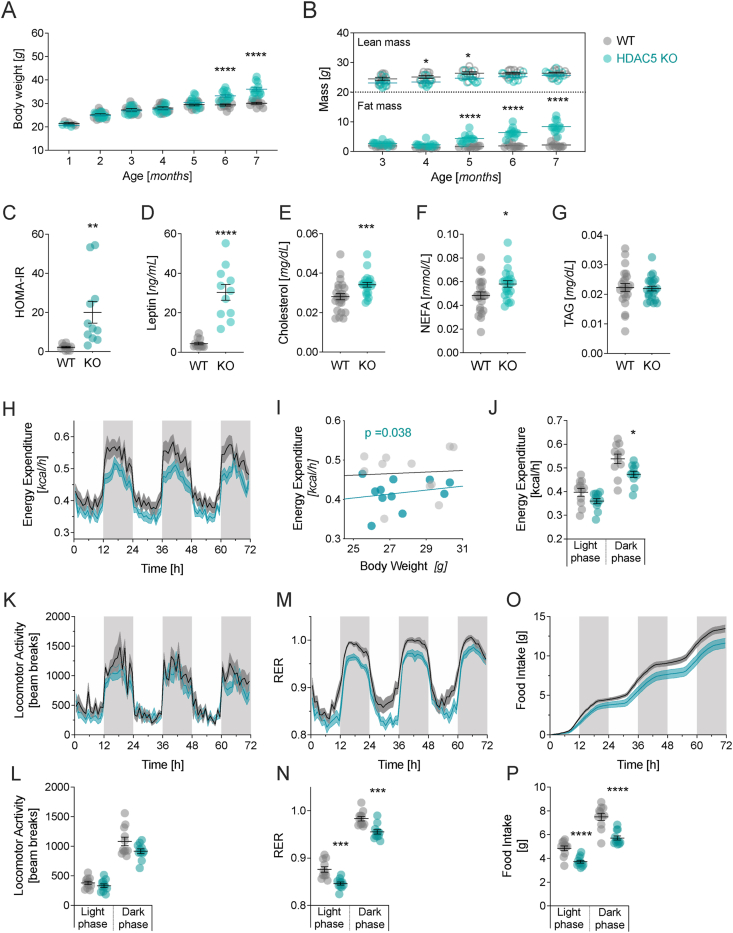
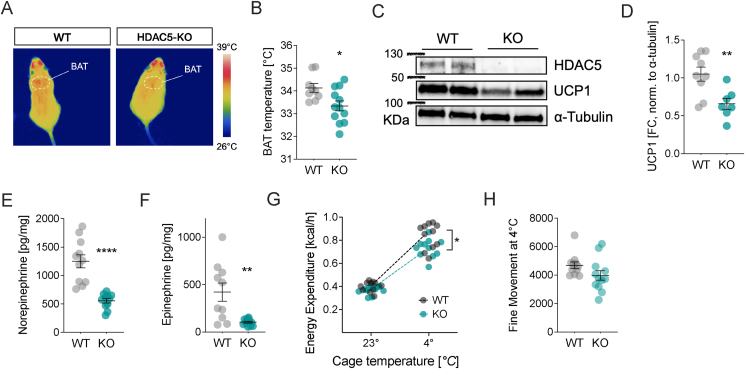
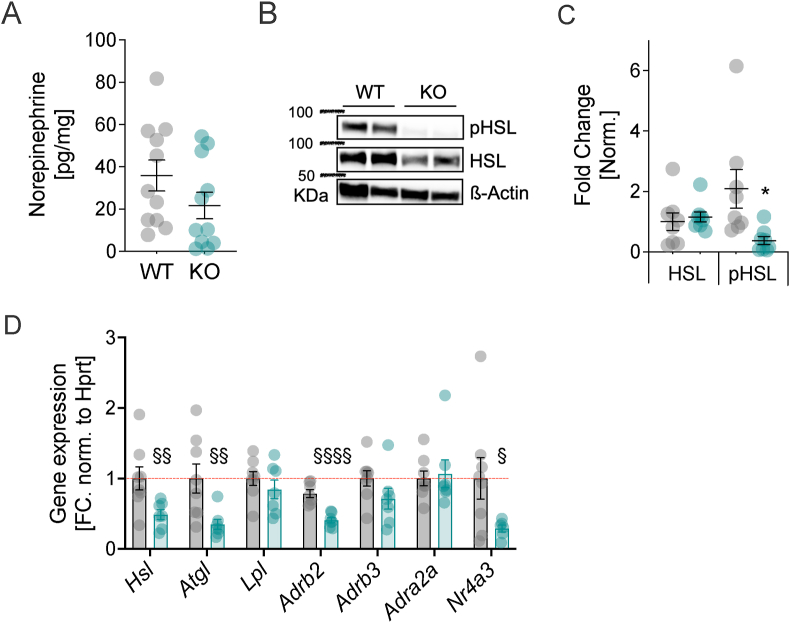
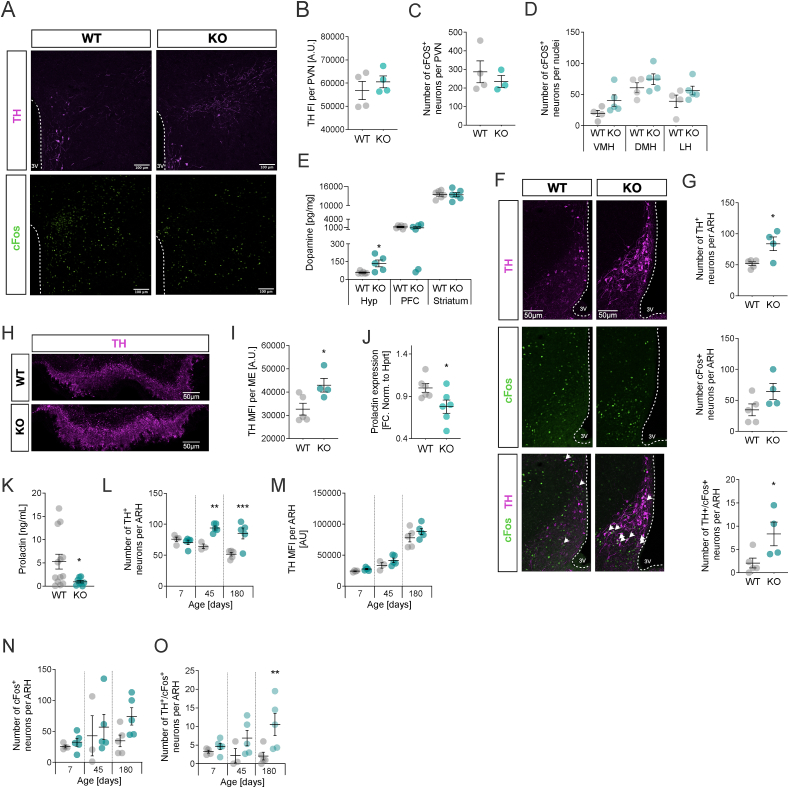
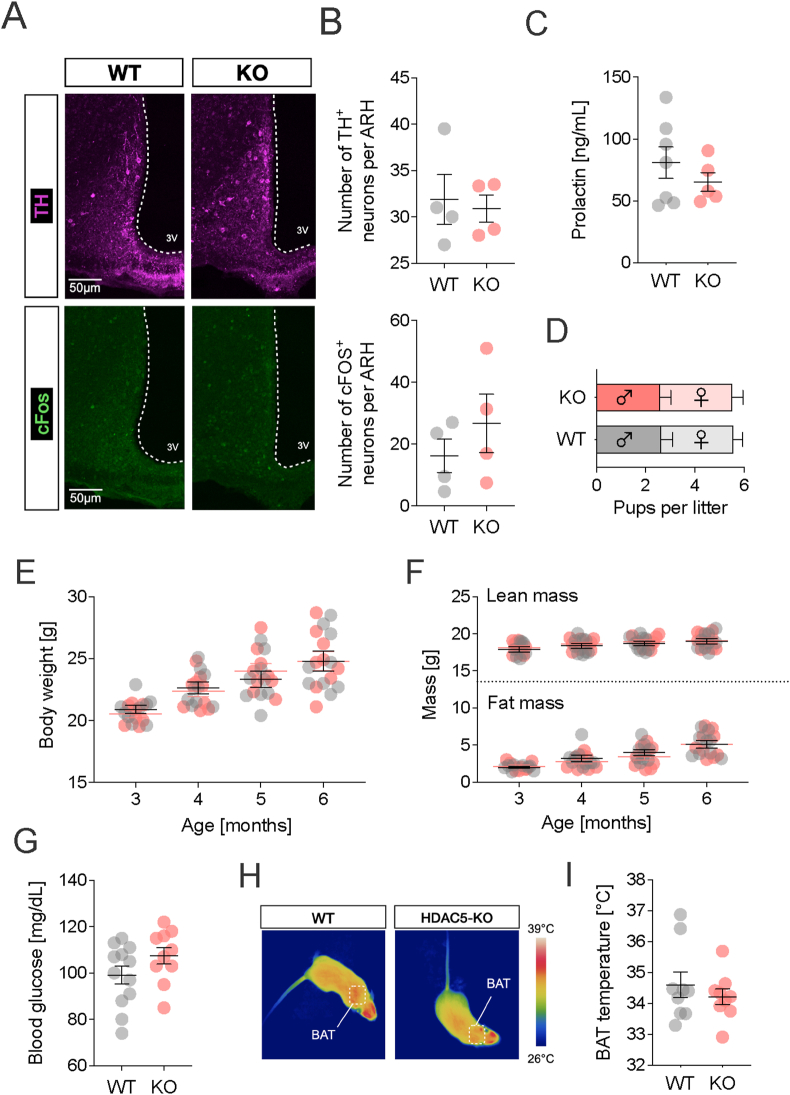
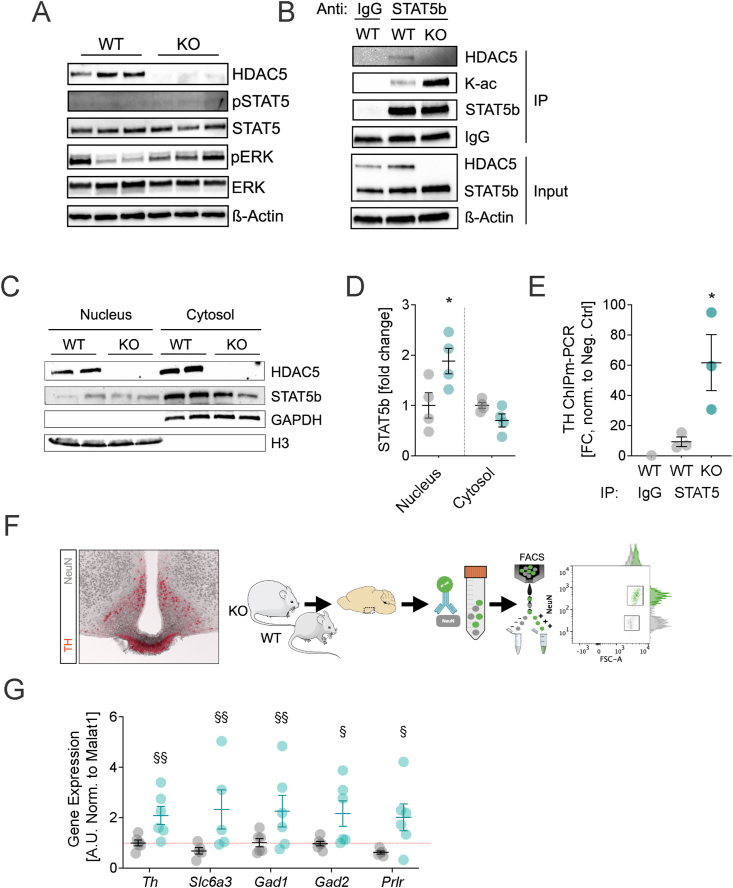
With age, metabolic perturbations accumulate to elevate our obesity burden. While age-onset obesity is mostly driven by a sedentary lifestyle and high calorie intake, genetic and epigenetic factors also play a role. Among these, members of the large histone deacetylase (HDAC) family are of particular importance as key metabolic determinants for healthy ageing, or metabolic dysfunction. Here, we aimed to interrogate the role of class 2 family member HDAC5 in controlling systemic metabolism and age-related obesity under non-obesogenic conditions. Starting at 6 months of age, we observed adult-onset obesity in chow-fed male global HDAC5-KO mice, that was accompanied by marked reductions in adrenergic-stimulated ATP-consuming futile cycles, including BAT activity and UCP1 levels, WAT-lipolysis, skeletal muscle, WAT and liver futile creatine and calcium cycles, and ultimately energy expenditure. Female mice did not differ between genotypes. The lower peripheral sympathetic nervous system (SNS) activity in mature male KO mice was linked to higher dopaminergic neuronal activity within the dorsomedial arcuate nucleus (dmARC) and elevated hypothalamic dopamine levels. Mechanistically, we reveal that hypothalamic HDAC5 acts as co-repressor of STAT5b over the control of Tyrosine hydroxylase (TH) gene transactivation, which ultimately orchestrates the activity of dmARH dopaminergic neurons and energy metabolism in male mice under non-obesogenic conditions.
Keywords: Adult-onset obesity; Brown fat thermogenesis; Dopamine; Futile ATP-consuming cycles; Histone deacetylase 5; Hypothalamus.
Copyright © 2024 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Conflict of interest statement
Declaration of competing interest The author is an Editorial Board Member/Editor-in-Chief/Associate Editor/Guest Editor for Molecular Metabolism and was not involved in the editorial review or the decision to publish this article. The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: PTP received speaker honoraria by Novo Nordisk. As a scientist, MHT participated in a scientific advisory board meeting of ERX Pharmaceuticals, Inc., Cambridge, MA, in 2019. He was a member of the Research Cluster Advisory Panel (ReCAP) of the Novo Nordisk Foundation between 2017 and 2019. He received funding for his research projects by Novo Nordisk (2016–2020) and Sanofi-Aventis (2012–2019). He consulted twice for Böhringer Ingelheim Pharma GmbH & Co. KG (2020 & 2021) and delivered a scientific lecture for Sanofi-Aventis Deutschland GmbH (2020) and for AstraZeneca GmbH (2024). As CEO and CSO of Helmholtz Munich, he is co-responsible for countless collaborations of the employees with a multitude of companies and institutions, worldwide. In this capacity, he discusses potential projects with and has signed/signs contracts for the centers institute(s) related to research collaborations worldwide, including but not limited to pharmaceutical corporations like Boehringer Ingelheim, Novo Nordisk, Roche Diagnostics, Arbormed, Eli Lilly, SCG Cell Therapy and others. As the CEO of Helmholtz Munich, he was/is further overall responsible for commercial technology transfer activities. MHT confirms that to the best of his knowledge none of the above funding sources or collaborations were involved in or had an influence on the preparation of this manuscript. All other authors declare that they have no conflict of interest related to this study.
Figures
References
-
- Fryar Cheryl D., Carroll Margaret D., Afful Joseph. Prevalence of overweight, obesity, and severe obesity among children and adolescents aged 2–19 Years: United States, 1963–1965 through 2017–2018. 2021. https://www.cdc.gov/nchs/data/hestat/obesity-child-17-18/obesity-child.htm
-
- Fryar Cheryl D., Carroll Margaret D., Afful Joseph. 2021. Prevalence of overweight, obesity, and extreme obesity among adults aged 20 and over: United States, 1960–1962 through 2017–2018.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
