

The Arabidopsis U1 snRNP regulates mRNA 3'-end processing
- PMID: 39313562
- PMCID: PMC11489095
- DOI: 10.1038/s41477-024-01796-8
The Arabidopsis U1 snRNP regulates mRNA 3'-end processing
Abstract
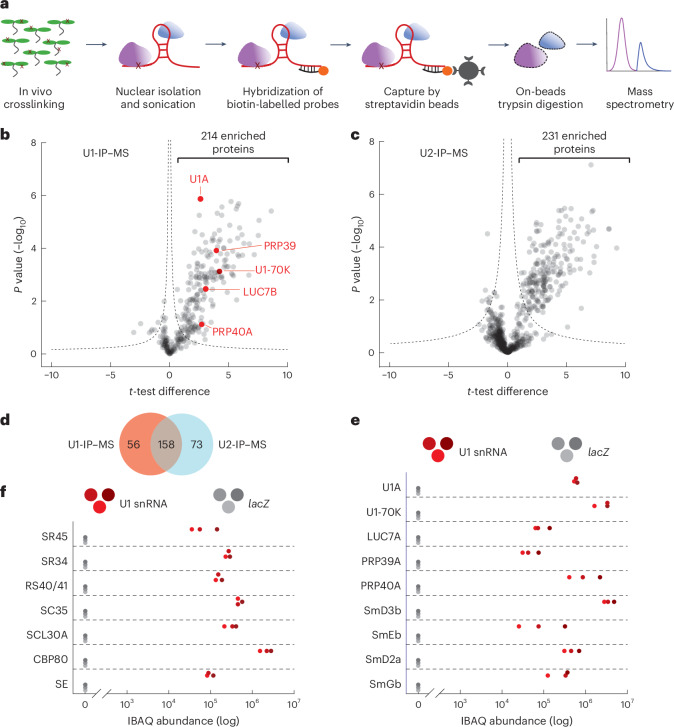
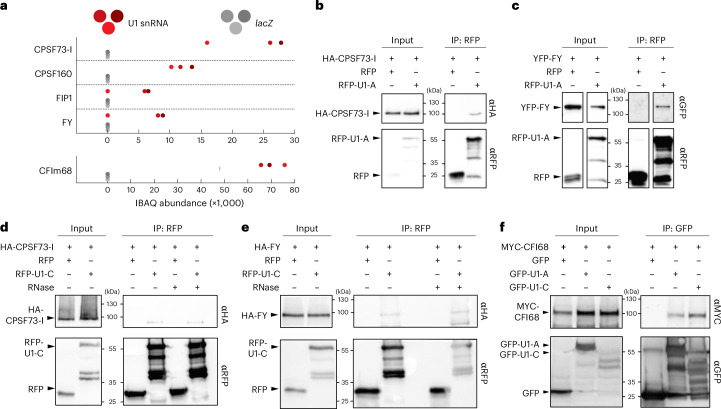
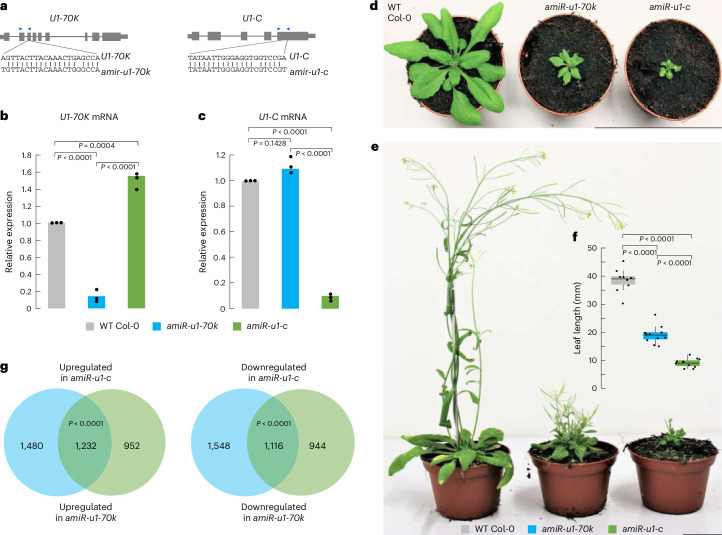
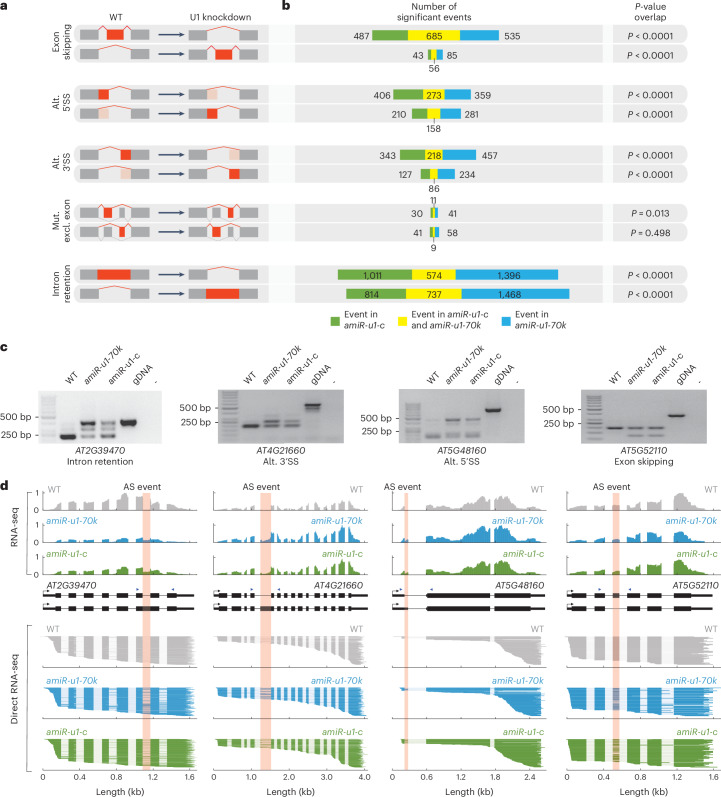
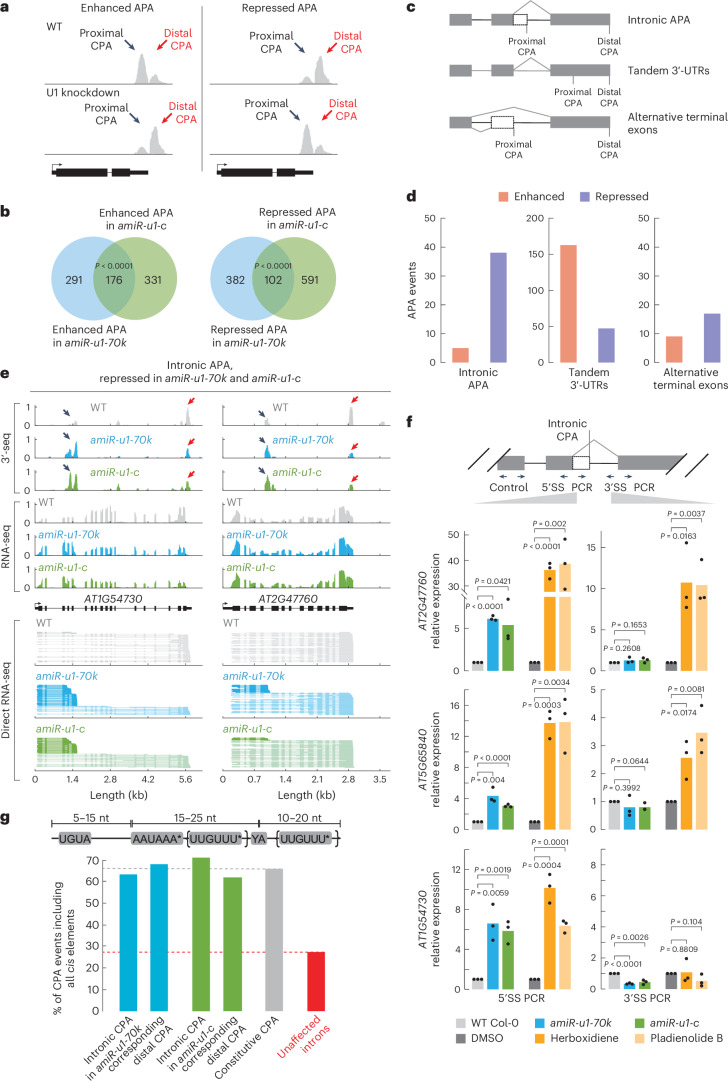
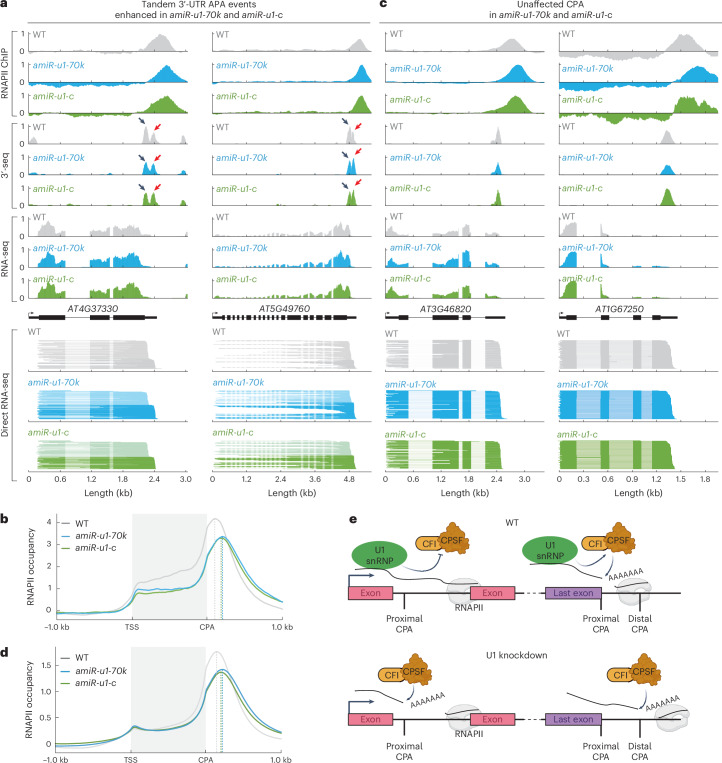
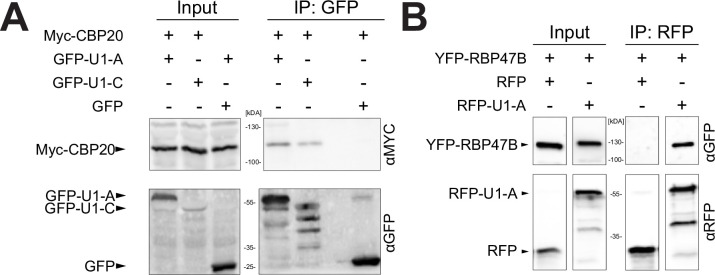
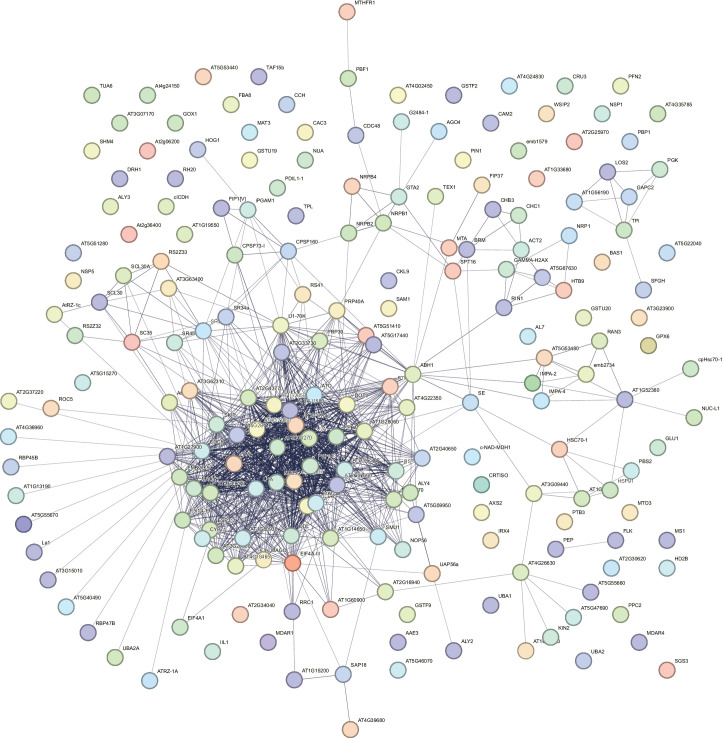
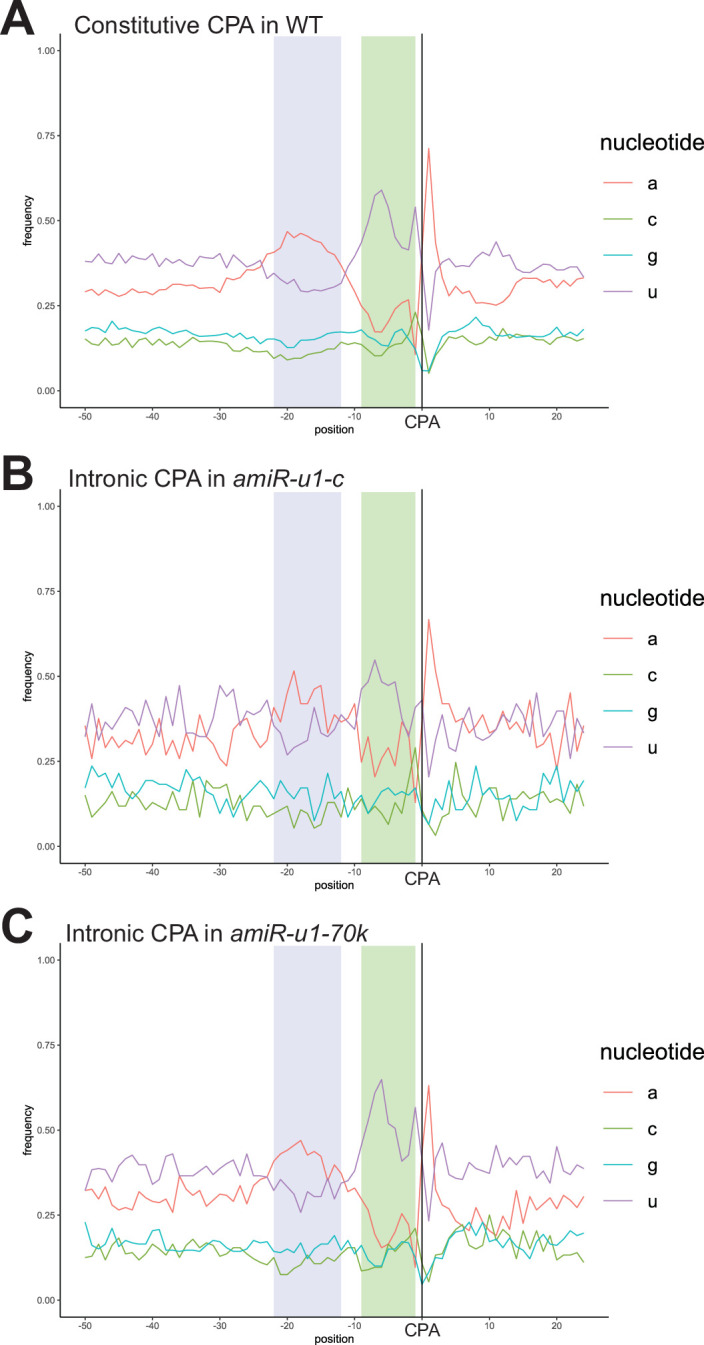
The removal of introns by the spliceosome is a key gene regulatory mechanism in eukaryotes, with the U1 snRNP subunit playing a crucial role in the early stages of splicing. Studies in metazoans show that the U1 snRNP also conducts splicing-independent functions, but the lack of genetic tools and knowledge about U1 snRNP-associated proteins have limited the study of such splicing-independent functions in plants. Here we describe an RNA-centric approach that identified more than 200 proteins associated with the Arabidopsis U1 snRNP and revealed a tight link to mRNA cleavage and polyadenylation factors. Interestingly, we found that the U1 snRNP protects mRNAs against premature cleavage and polyadenylation within introns-a mechanism known as telescripting in metazoans-while also influencing alternative polyadenylation site selection in 3'-UTRs. Overall, our work provides a comprehensive view of U1 snRNP interactors and reveals novel functions in regulating mRNA 3'-end processing in Arabidopsis, laying the groundwork for understanding non-canonical functions of plant U1 snRNPs.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
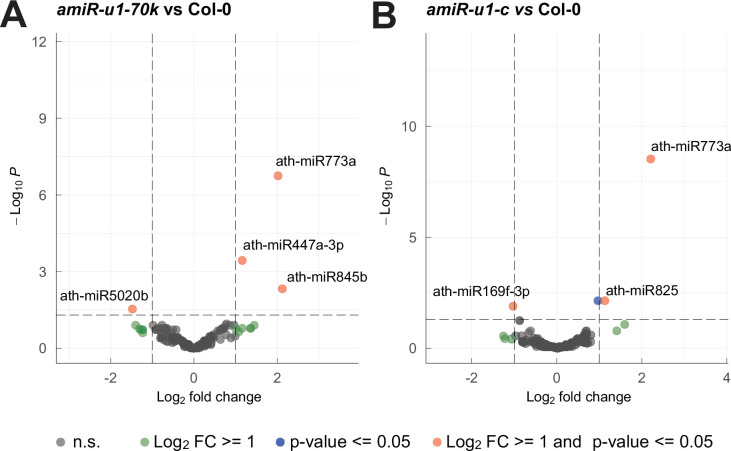
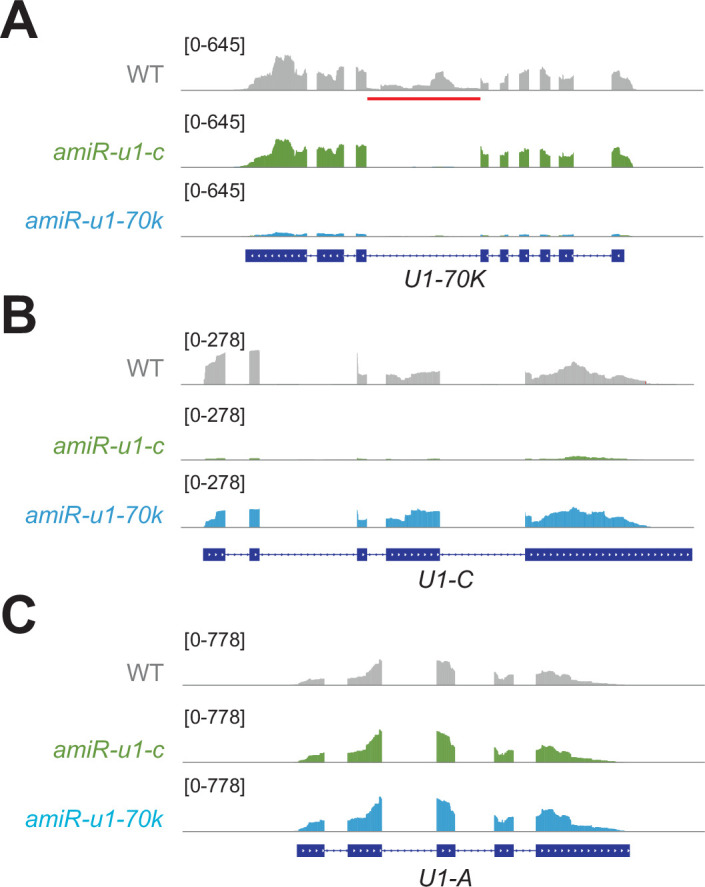
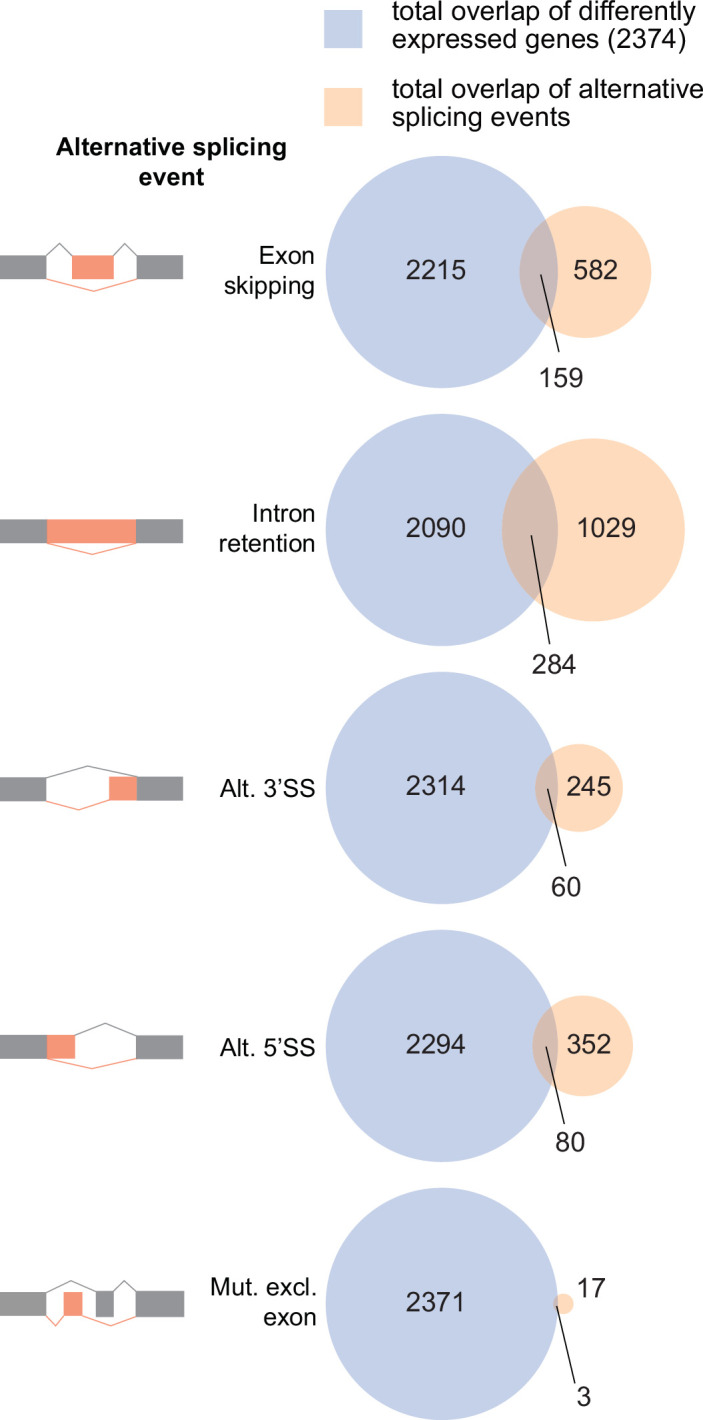
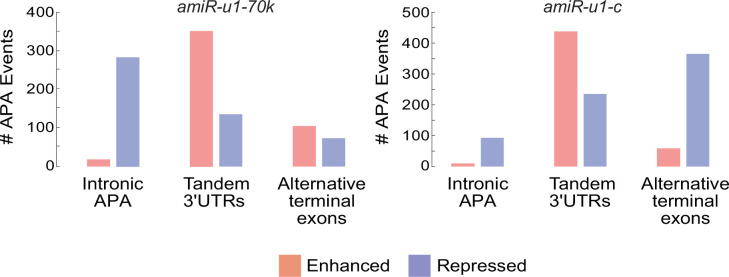
References
-
- Chen, W. & Moore, M. J. Spliceosomes. Curr. Biol.25, R181–R183 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
