

This is a preprint.
"Mitotic" kinesin-5 is a dynamic brake for axonal growth
- PMID: 39314406
- PMCID: PMC11419024
- DOI: 10.1101/2024.09.12.612721
"Mitotic" kinesin-5 is a dynamic brake for axonal growth
Update in
-
'Mitotic' kinesin-5 is a dynamic brake for axonal growth in Drosophila.Development. 2025 May 1;152(9):dev204424. doi: 10.1242/dev.204424. Epub 2025 May 8. Development. 2025. PMID: 40223510
Abstract
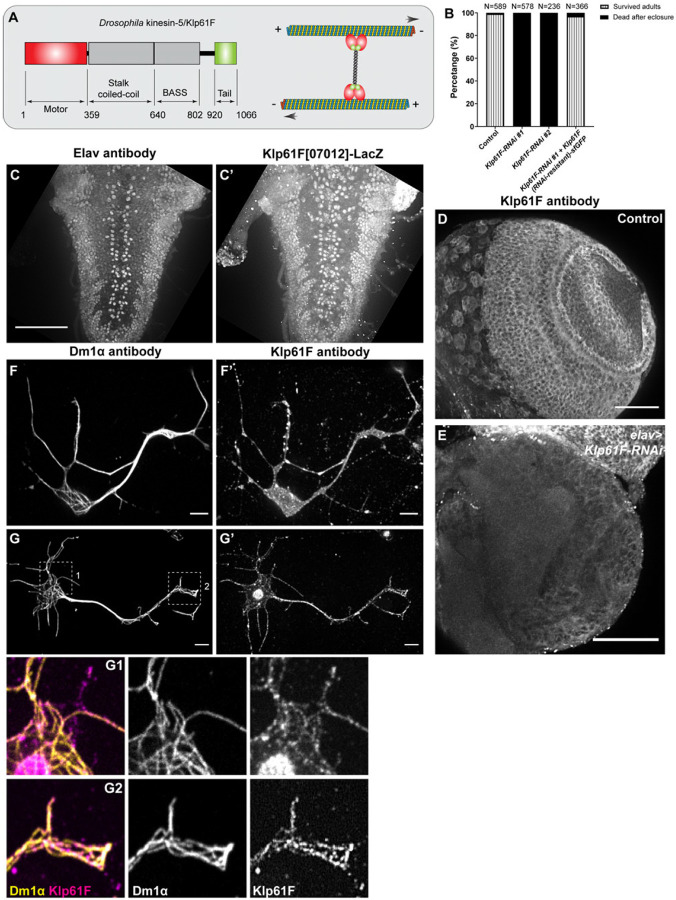
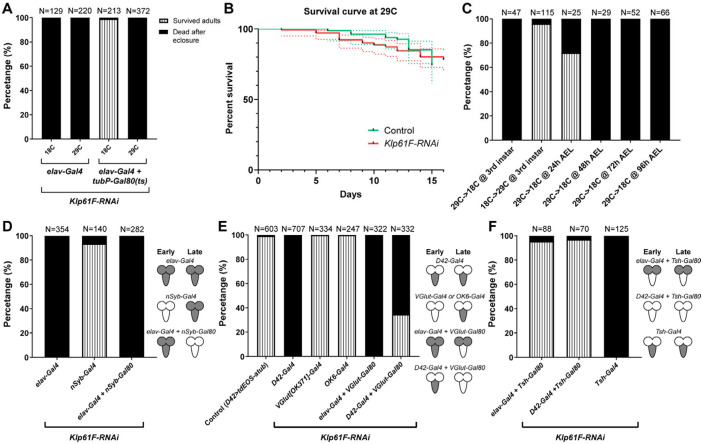
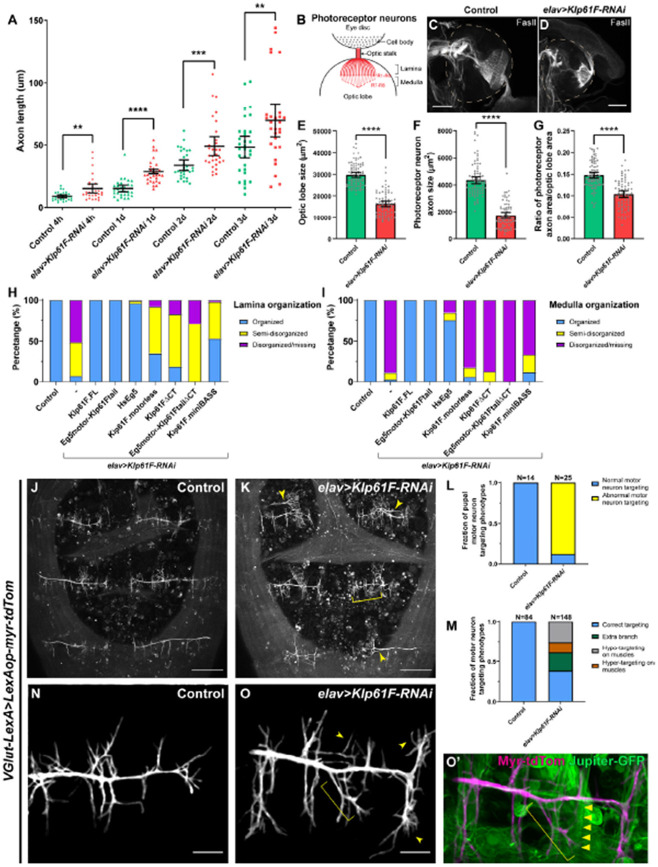
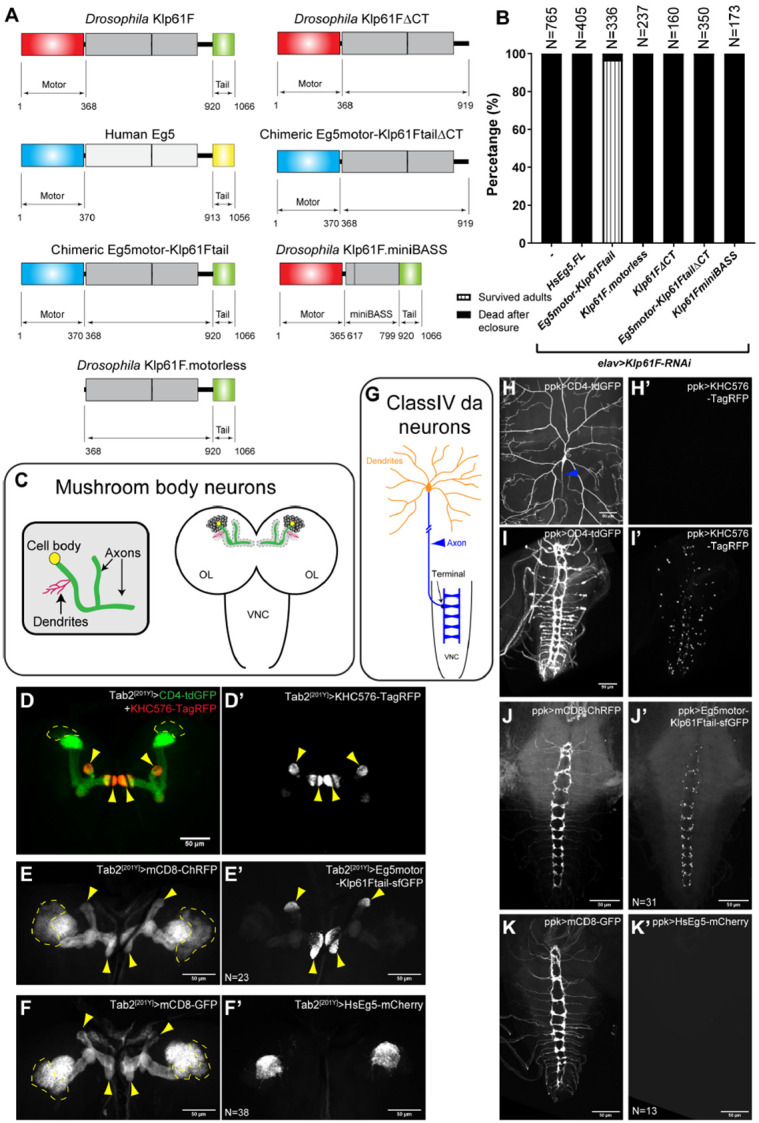
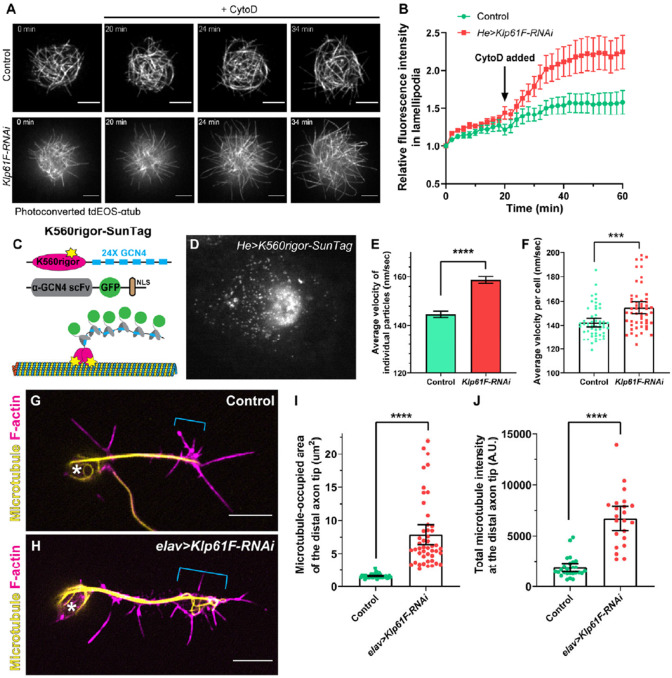
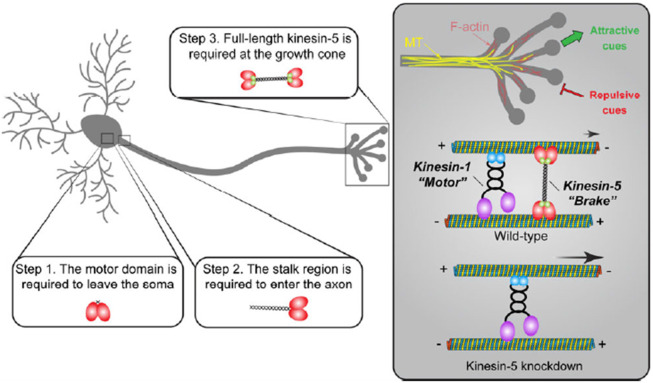
During neuronal development, neurons undergo significant microtubule reorganization to shape axons and dendrites, establishing the framework for efficient wiring of the nervous system. Previous studies from our laboratory demonstrated the key role of kinesin-1 in driving microtubule-microtubule sliding, which provides the mechanical forces necessary for early axon outgrowth and regeneration in Drosophila melanogaster. In this study, we reveal the critical role of kinesin-5, a mitotic motor, in modulating the development of postmitotic neurons. Kinesin-5, a conserved homotetrameric motor, typically functions in mitosis by sliding antiparallel microtubules apart in the spindle. Here, we demonstrate that the Drosophila kinesin-5 homolog, Klp61F, is expressed in larval brain neurons, with high levels in ventral nerve cord (VNC) neurons. Knockdown of Klp61F using a pan-neuronal driver leads to severe locomotion defects and complete lethality in adult flies, mainly due to the absence of kinesin-5 in VNC motor neurons during early larval development. Klp61F depletion results in significant axon growth defects, both in cultured and in vivo neurons. By imaging individual microtubules, we observe a significant increase in microtubule motility, and excessive penetration of microtubules into the axon growth cone in Klp61F-depleted neurons. Adult lethality and axon growth defects are fully rescued by a chimeric human-Drosophila kinesin-5 motor, which accumulates at the axon tips, suggesting a conserved role of kinesin-5 in neuronal development. Altogether, our findings show that at the growth cone, kinesin-5 acts as a brake on kinesin-1-driven microtubule sliding, preventing premature microtubule entry into the growth cone. This regulatory role of kinesin-5 is essential for precise axon pathfinding during nervous system development.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources