

This is a preprint.
Nutrient starvation activates ECM remodeling gene enhancers associated with inflammatory bowel disease risk in fibroblasts
- PMID: 39314475
- PMCID: PMC11418948
- DOI: 10.1101/2024.09.06.611754
Nutrient starvation activates ECM remodeling gene enhancers associated with inflammatory bowel disease risk in fibroblasts
Abstract
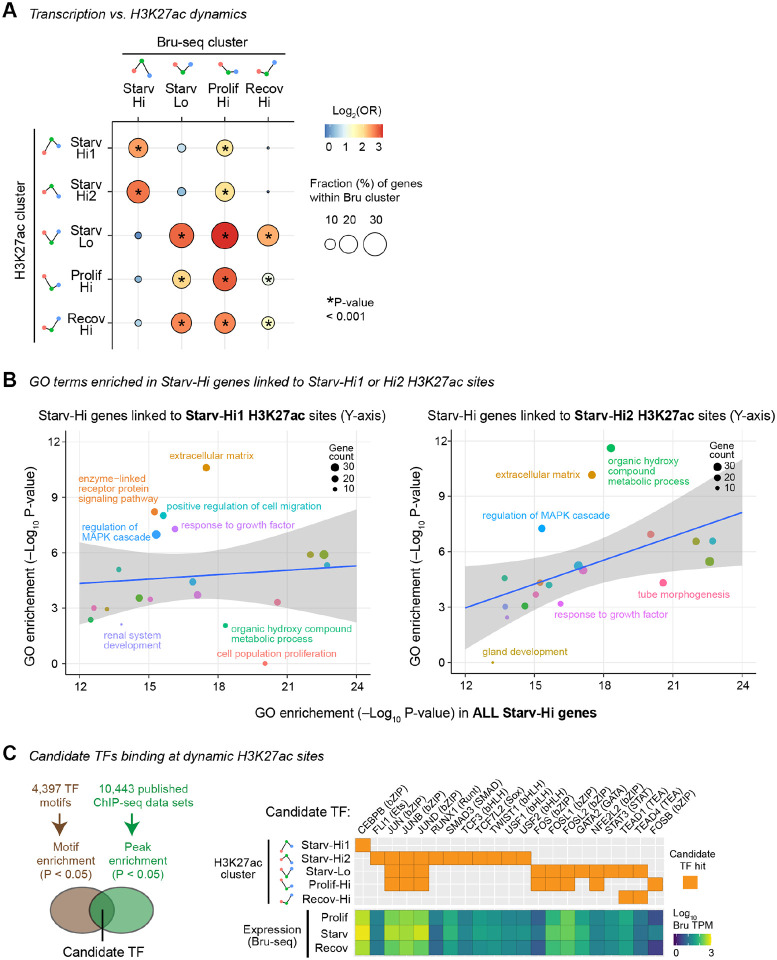
Nutrient deprivation induces a reversible cell cycle arrest state termed quiescence, which often accompanies transcriptional silencing and chromatin compaction. Paradoxically, nutrient deprivation is associated with activated fibroblast states in pathological microenvironments in which fibroblasts drive extracellular matrix (ECM) remodeling to alter tissue environments. The relationship between nutrient deprivation and fibroblast activation remains unclear. Here, we report that serum deprivation extensively activates transcription of ECM remodeling genes in cultured fibroblasts, despite the induction of quiescence. Starvation-induced transcriptional activation accompanied large-scale histone acetylation of putative distal enhancers, but not promoters. The starvation-activated putative enhancers were enriched for non-coding genetic risk variants associated with inflammatory bowel disease (IBD), suggesting that the starvation-activated gene regulatory network may contribute to fibroblast activation in IBD. Indeed, the starvation-activated gene PLAU, encoding uPA serine protease for plasminogen and ECM, was upregulated in inflammatory fibroblasts in the intestines of IBD patients. Furthermore, the starvation-activated putative enhancer at PLAU, which harbors an IBD risk variant, gained chromatin accessibility in IBD patient fibroblasts. This study implicates nutrient deprivation in transcriptional activation of ECM remodeling genes in fibroblasts and suggests nutrient deprivation as a potential mechanism for pathological fibroblast activation in IBD.
Conflict of interest statement
DECLARATION OF INTERESTS The authors declare no competing interests.
Figures




Similar articles
-
Starvation activates ECM-remodeling gene transcription and putative enhancers in fibroblasts despite inducing quiescence.Cell Rep. 2025 Jul 22;44(7):115896. doi: 10.1016/j.celrep.2025.115896. Epub 2025 Jun 24. Cell Rep. 2025. PMID: 40560730 Free PMC article.
-
Inflammatory Bowel Disease-Associated Gut Commensals Degrade Components of the Extracellular Matrix.mBio. 2022 Dec 20;13(6):e0220122. doi: 10.1128/mbio.02201-22. Epub 2022 Nov 29. mBio. 2022. PMID: 36445085 Free PMC article.
-
Hyaluronan synthase-2 upregulation protects smpd3-deficient fibroblasts against cell death induced by nutrient deprivation, but not against apoptosis evoked by oxidized LDL.Redox Biol. 2015;4:118-26. doi: 10.1016/j.redox.2014.12.004. Epub 2014 Dec 16. Redox Biol. 2015. PMID: 25555205 Free PMC article.
-
The intestinal tissue homeostasis - the role of extracellular matrix remodeling in inflammatory bowel disease.Expert Rev Gastroenterol Hepatol. 2019 Oct;13(10):977-993. doi: 10.1080/17474124.2019.1673729. Epub 2019 Oct 22. Expert Rev Gastroenterol Hepatol. 2019. PMID: 31587588 Review.
-
The extracellular matrix in IBD: a dynamic mediator of inflammation.Curr Opin Gastroenterol. 2017 Jul;33(4):234-238. doi: 10.1097/MOG.0000000000000368. Curr Opin Gastroenterol. 2017. PMID: 28562487 Free PMC article. Review.
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous