

This is a preprint.
A secondary β-hydroxybutyrate metabolic pathway linked to energy balance
- PMID: 39314488
- PMCID: PMC11418978
- DOI: 10.1101/2024.09.09.612087
A secondary β-hydroxybutyrate metabolic pathway linked to energy balance
Abstract
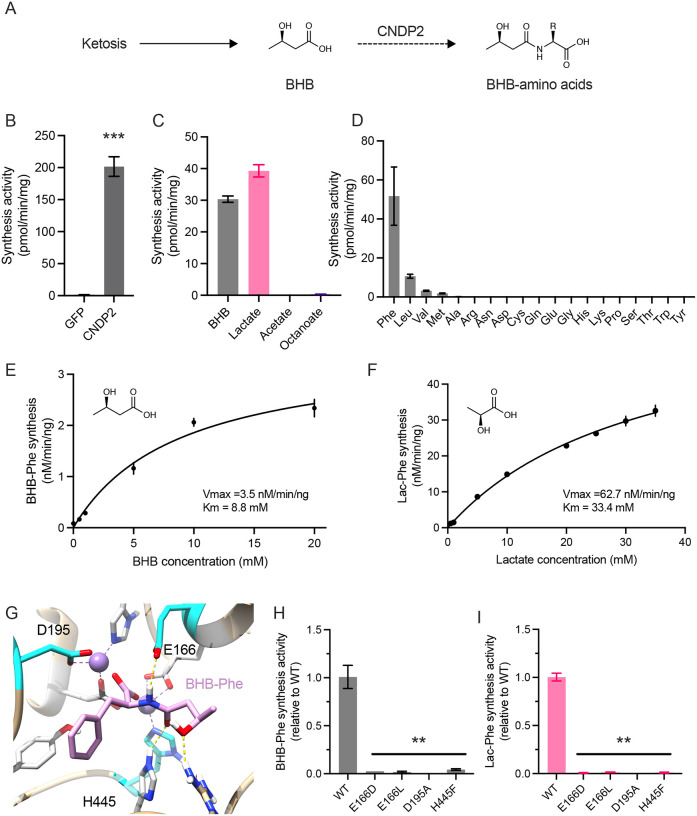
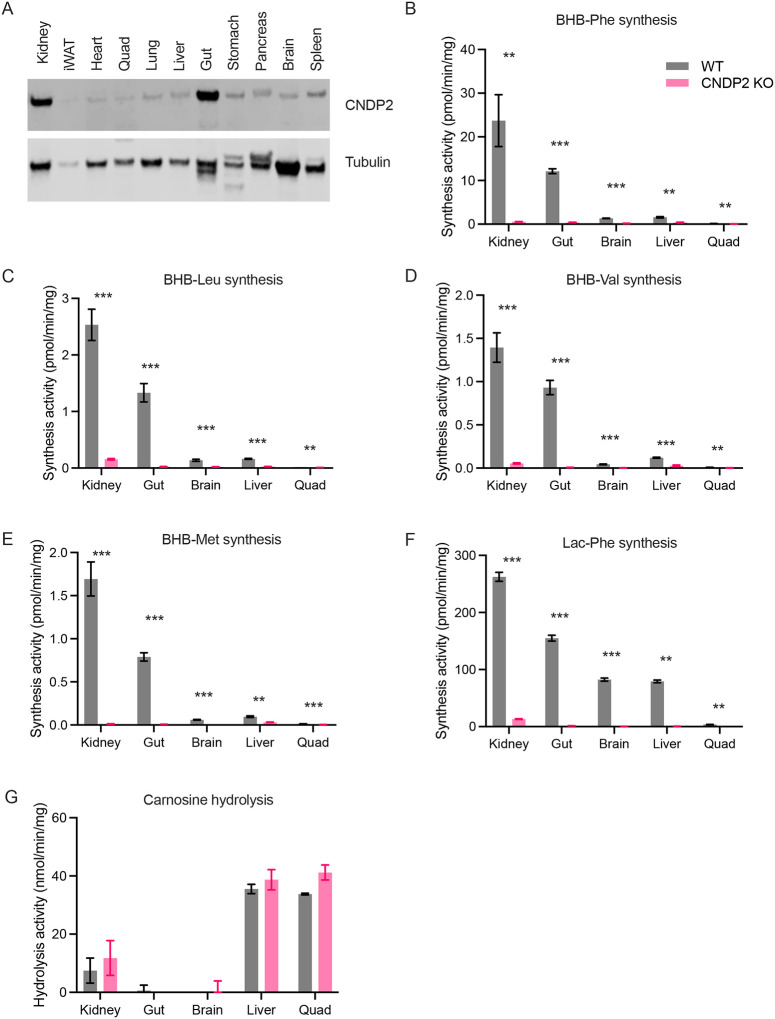
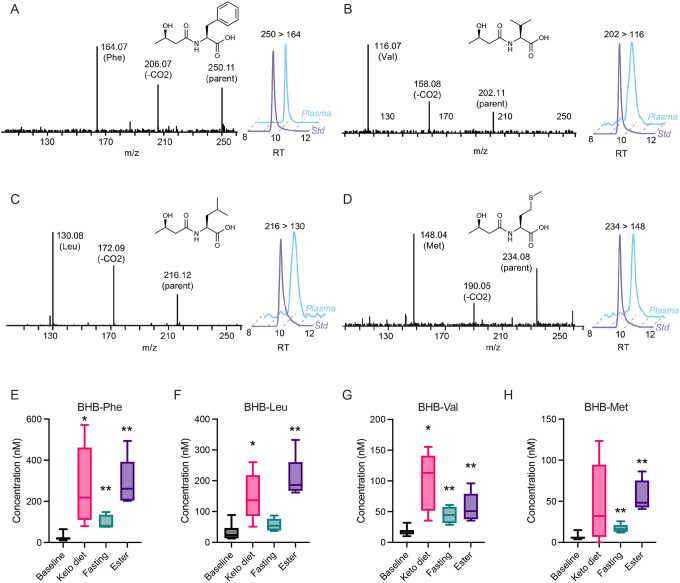
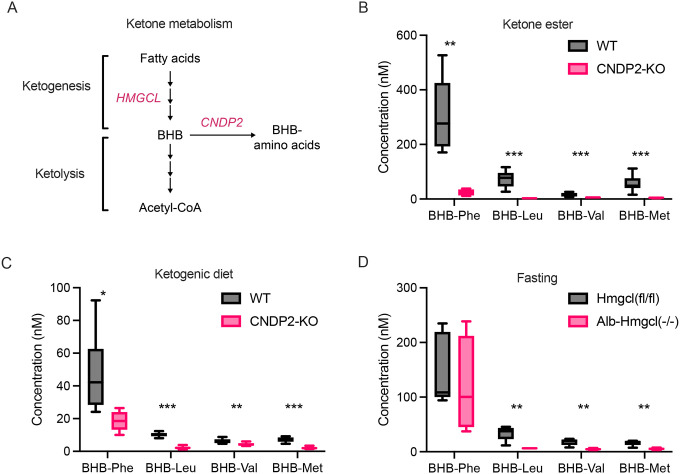
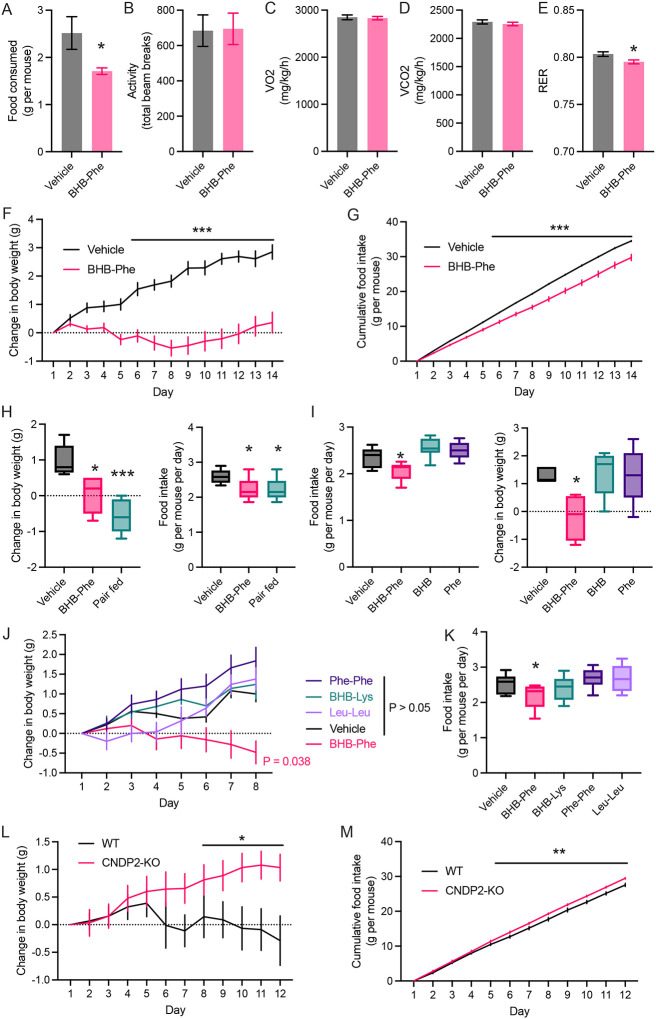
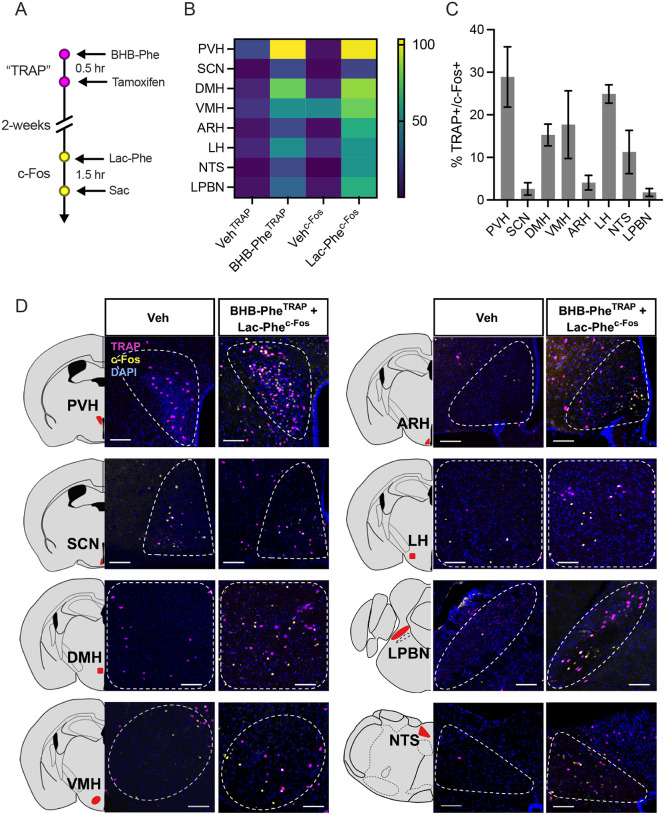
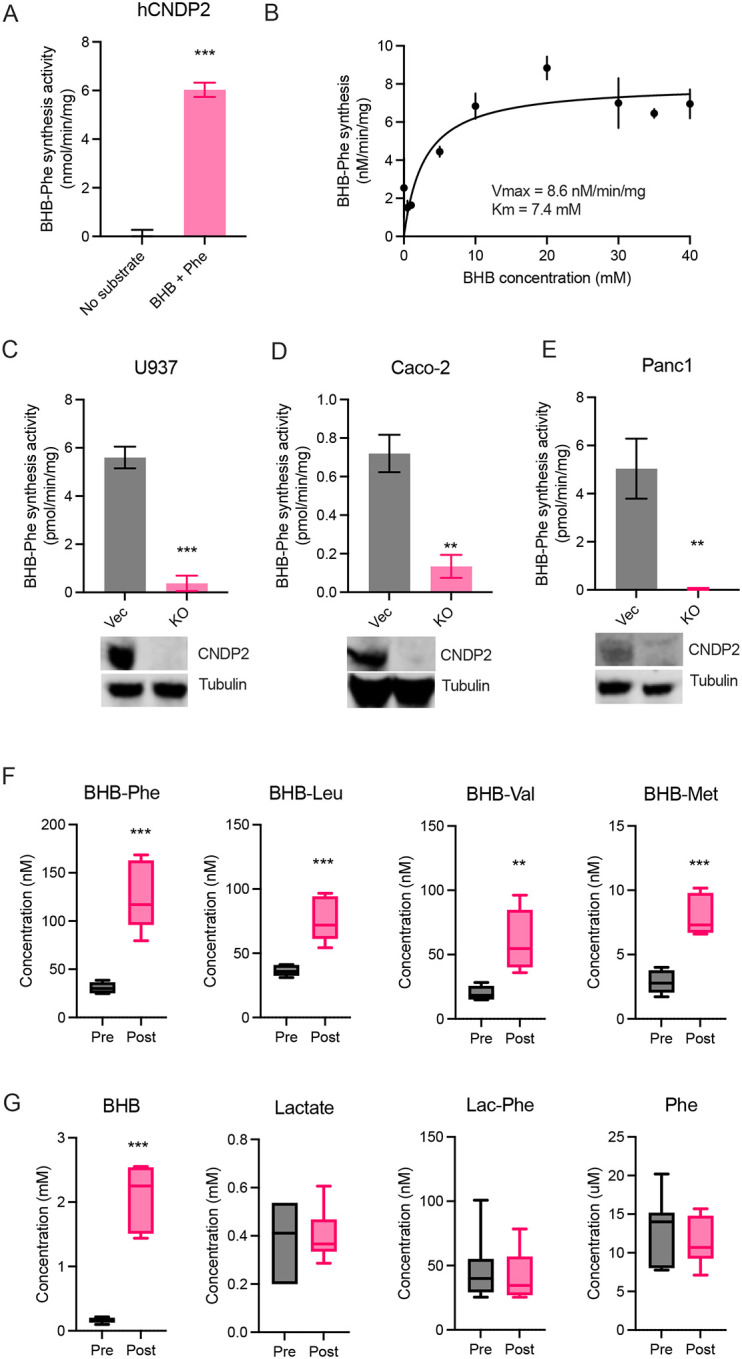
β-hydroxybutyrate (BHB) is an abundant ketone body. To date, all known pathways of BHB metabolism involve interconversion of BHB and primary energy intermediates. Here we show that CNDP2 controls a previously undescribed secondary BHB metabolic pathway via enzymatic conjugation of BHB and free amino acids. This BHB-ylation reaction produces a family of endogenous ketone metabolites, the BHB-amino acids. Genetic ablation of CNDP2 in mice eliminates tissue amino acid BHB-ylation activity and reduces BHB-amino acid levels. Administration of BHB-Phe, the most abundant BHB-amino acid, to obese mice activates neural populations in the hypothalamus and brainstem and suppresses feeding and body weight. Conversely, CNDP2-KO mice exhibit increased food intake and body weight upon ketosis stimuli. CNDP2-dependent amino acid BHB-ylation and BHB-amino acid metabolites are also conserved in humans. Therefore, the metabolic pathways of BHB extend beyond primary metabolism and include secondary ketone metabolites linked to energy balance.
Conflict of interest statement
Declaration of interests A provisional patent application has been filed by Stanford University on BHB-amino acids for the treatment of cardiometabolic disease.
Figures
Similar articles
-
A β-hydroxybutyrate shunt pathway generates anti-obesity ketone metabolites.Cell. 2025 Jan 9;188(1):175-186.e20. doi: 10.1016/j.cell.2024.10.032. Epub 2024 Nov 12. Cell. 2025. PMID: 39536746
-
Beta-Hydroxybutyrate: A Supplemental Molecule for Various Diseases.Curr Drug Targets. 2024;25(14):919-933. doi: 10.2174/0113894501312168240821082224. Curr Drug Targets. 2024. PMID: 39238395 Review.
-
Genetic and nongenetic profiling of milk β-hydroxybutyrate and acetone and their associations with ketosis in Holstein cows.J Dairy Sci. 2020 Nov;103(11):10332-10346. doi: 10.3168/jds.2020-18339. Epub 2020 Sep 18. J Dairy Sci. 2020. PMID: 32952022
-
The ketone body β-hydroxybutyrate alleviates CoCrMo alloy particles induced osteolysis by regulating NLRP3 inflammasome and osteoclast differentiation.J Nanobiotechnology. 2022 Mar 9;20(1):120. doi: 10.1186/s12951-022-01320-0. J Nanobiotechnology. 2022. PMID: 35264201 Free PMC article.
-
A systematic review and meta-regression of exogenous ketone infusion rates and resulting ketosis-A tool for clinicians and researchers.Front Physiol. 2023 Jun 28;14:1202186. doi: 10.3389/fphys.2023.1202186. eCollection 2023. Front Physiol. 2023. PMID: 37449016 Free PMC article.
References
-
- Rahman M. et al. The b-hydroxybutyrate receptor HCA 2 activates a neuroprotective subset of macrophages. Nat. Commun. 5, 1–11 (2014). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous