

Update on a brain-penetrant cardiac glycoside that can lower cellular prion protein levels in human and guinea pig paradigms
- PMID: 39316592
- PMCID: PMC11421771
- DOI: 10.1371/journal.pone.0308821
Update on a brain-penetrant cardiac glycoside that can lower cellular prion protein levels in human and guinea pig paradigms
Abstract
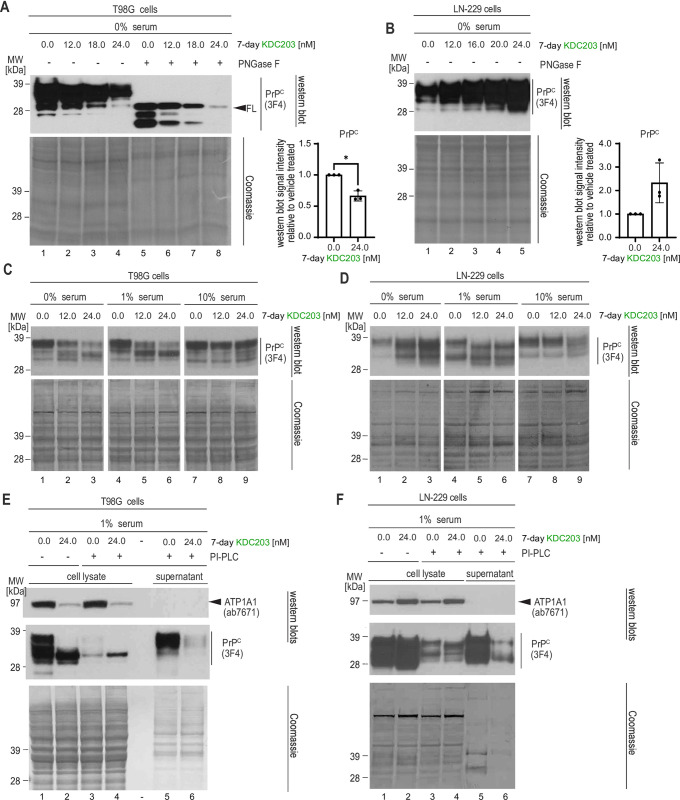
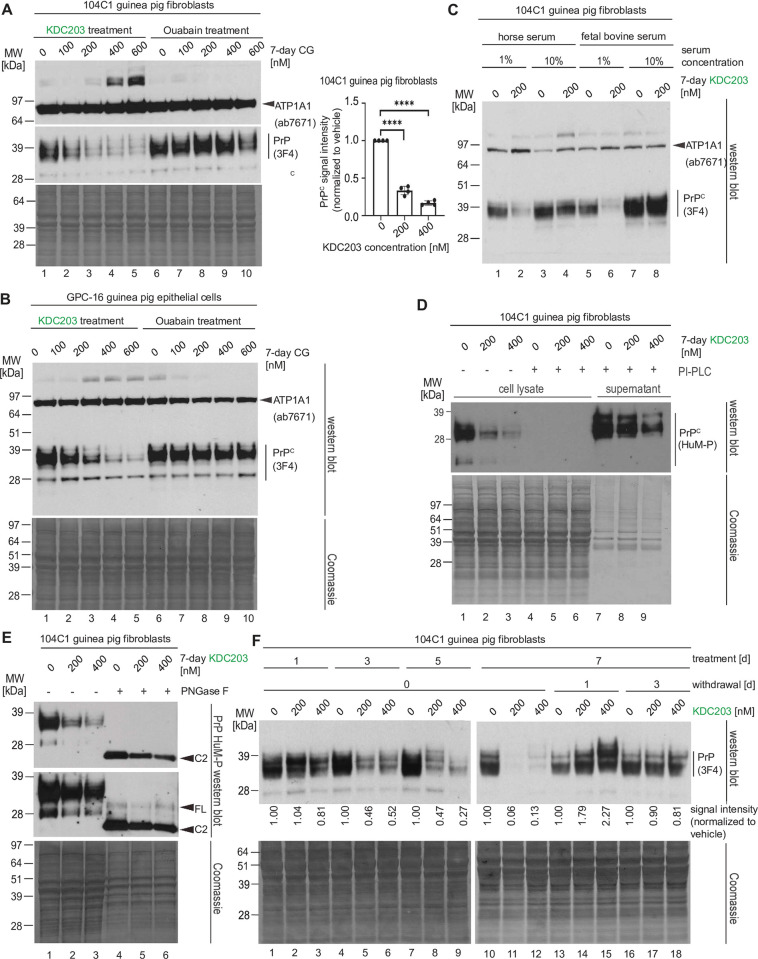
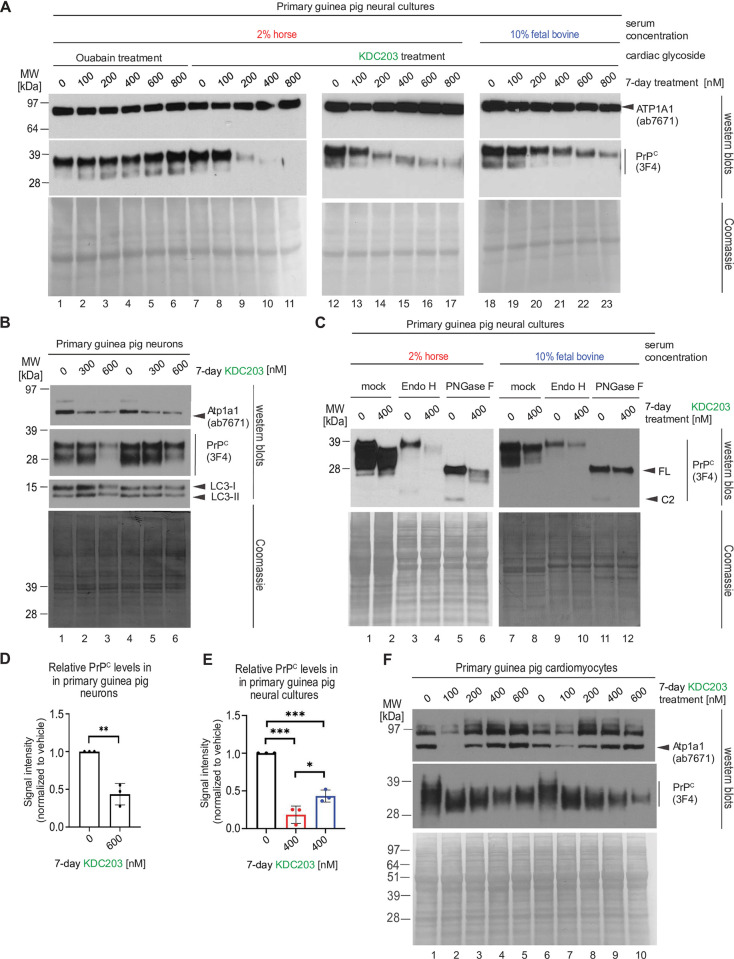
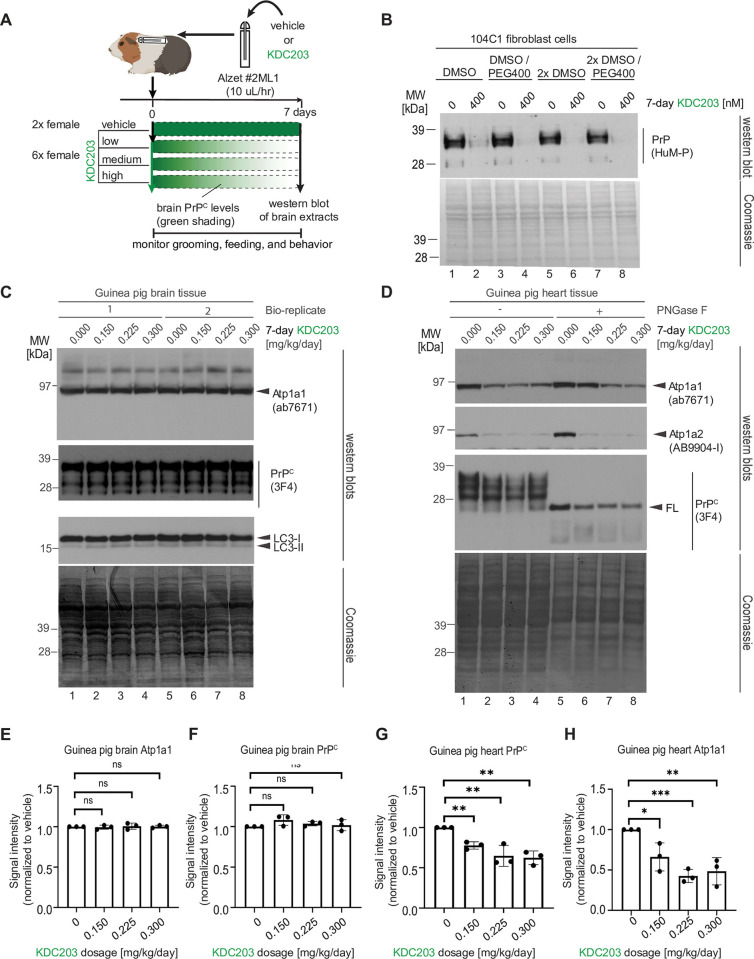
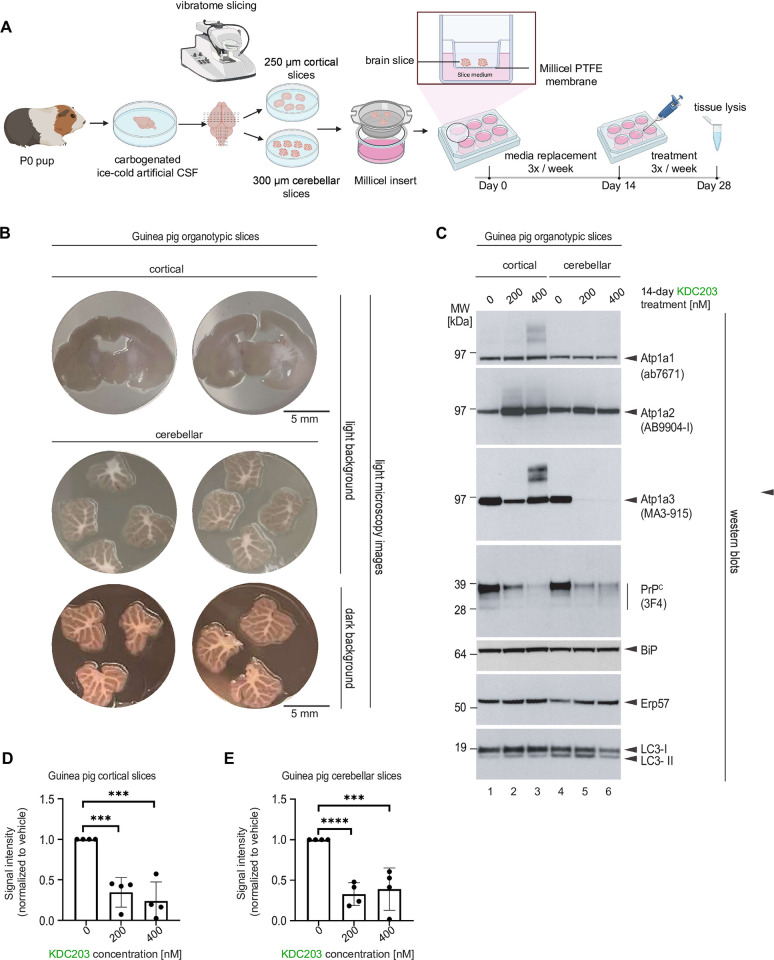
Lowering the levels of the cellular prion protein (PrPC) is widely considered a promising strategy for the treatment of prion diseases. Building on work that established immediate spatial proximity of PrPC and Na+, K+-ATPases (NKAs) in the brain, we recently showed that PrPC levels can be reduced by targeting NKAs with their natural cardiac glycoside (CG) inhibitors. We then introduced C4'-dehydro-oleandrin as a CG with improved pharmacological properties for this indication, showing that it reduced PrPC levels by 84% in immortalized human cells that had been differentiated to acquire neural or astrocytic characteristics. Here we report that our lead compound caused cell surface PrPC levels to drop also in other human cell models, even when the analyses of whole cell lysates suggested otherwise. Because mice are refractory to CGs, we explored guinea pigs as an alternative rodent model for the preclinical evaluation of C4'-dehydro-oleandrin. We found that guinea pig cell lines, primary cells, and brain slices were responsive to our lead compound, albeit it at 30-fold higher concentrations than human cells. Of potential significance for other PrPC lowering approaches, we observed that cells attempted to compensate for the loss of cell surface PrPC levels by increasing the expression of the prion gene, requiring daily administration of C4'-dehydro-oleandrin for a sustained PrPC lowering effect. Regrettably, when administered systemically in vivo, the levels of C4'-dehydro-oleandrin that reached the guinea pig brain remained insufficient for the PrPC lowering effect to manifest. A more suitable preclinical model is still needed to determine if C4'-dehydro-oleandrin can offer a cost-effective complementary strategy for pushing PrPC levels below a threshold required for long-term prion disease survival.
Copyright: © 2024 Eid et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: GS and DW declare that they are co-inventors on patent applications named ‘Compounds for altering levels of one or more NKA alpha subunits and their use in treating prion diseases or brain diseases associated with cellular prion protein’ filed in the US and Canada as well as a European patent application named ‘Compounds and methods to treat prion and related diseases’ based on a PCT international filing (PCT/CA2022/050358). These patent applications describe the use of CGs for the purpose of reducing steady-state levels of PrPC. This potential conflict-of-interest does not affect the adherence of all authors to journal policies on the sharing of data and materials. This does not alter our adherence to PLoS One policies on sharing data and materials.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
