

Optimized Proteomic Analysis of Insulin Granules From MIN6 Cells Identifies Scamp3, a Novel Regulator of Insulin Secretion and Content
- PMID: 39320956
- PMCID: PMC11579411
- DOI: 10.2337/db24-0355
Optimized Proteomic Analysis of Insulin Granules From MIN6 Cells Identifies Scamp3, a Novel Regulator of Insulin Secretion and Content
Abstract
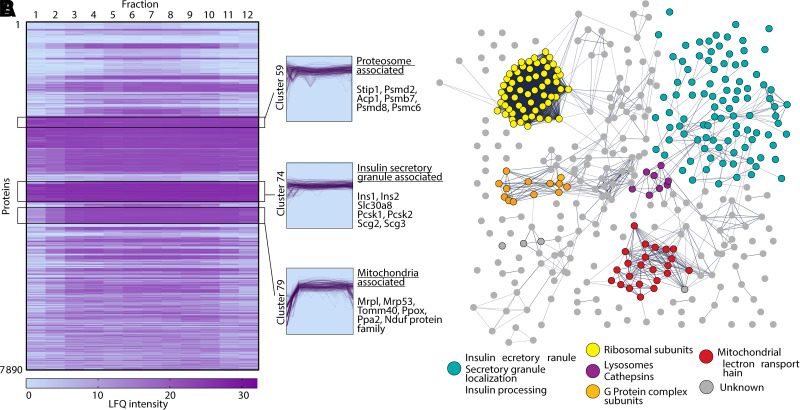
Pancreatic β-cells in the islets of Langerhans are key to maintaining glucose homeostasis by secreting the peptide hormone insulin. Insulin is packaged within vesicles named insulin secretory granules (ISGs), which recently have been considered to have intrinsic structures and proteins that regulate insulin granule maturation, trafficking, and secretion. Previously, studies have identified a handful of novel ISG-associated proteins, using different separation techniques. The present study combines an optimized ISG isolation technique and mass spectrometry-based proteomics, with an unbiased protein correlation profiling and targeted machine-learning approach to uncover 211 ISG-associated proteins with confidence. Four of these proteins, syntaxin-7, synaptophysin, synaptotagmin-13, and Scamp3 have not been previously associated with ISG. Through colocalization analysis of confocal imaging, we validate the association of these proteins to the ISG in MIN6 and human β-cells. We further validate the role for one (Scamp3) in regulating insulin content and secretion from β-cells for the first time. Scamp3 knockdown INS-1 cells have reduced insulin content and dysfunctional insulin secretion. These data provide the basis for future investigation of Scamp3 in β-cell biology and the regulation of insulin secretion.
© 2024 by the American Diabetes Association.
Conflict of interest statement
Figures




References
-
- Gandasi NR, Yin P, Omar-Hmeadi M, et al. Glucose-dependent granule docking limits insulin secretion and is decreased in human type 2 diabetes. Cell Metab 2018;27:470–478 e474 - PubMed
-
- Ramzy A, Asadi A, Kieffer TJ. Revisiting proinsulin processing: evidence that human β-cells process proinsulin with prohormone convertase (PC) 1/3 but not PC2. Diabetes 2020;69:1451–1462 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
