

ST8Sia2 polysialyltransferase protects against infection by Trypanosoma cruzi
- PMID: 39321148
- PMCID: PMC11466412
- DOI: 10.1371/journal.pntd.0012454
ST8Sia2 polysialyltransferase protects against infection by Trypanosoma cruzi
Abstract
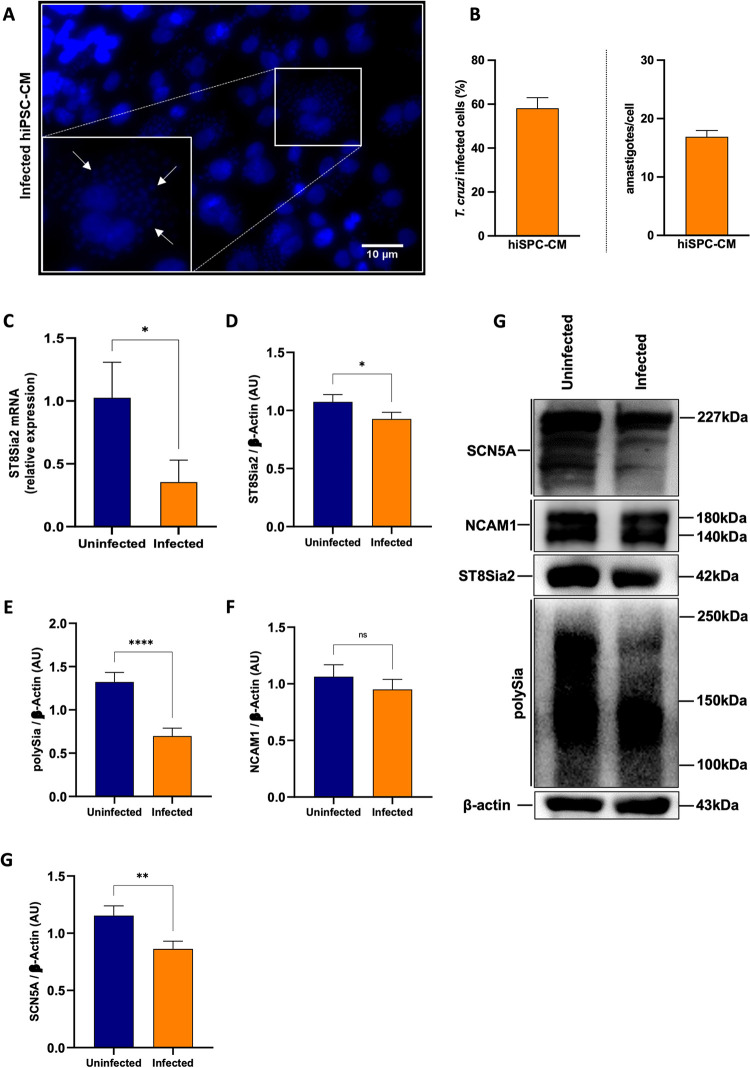
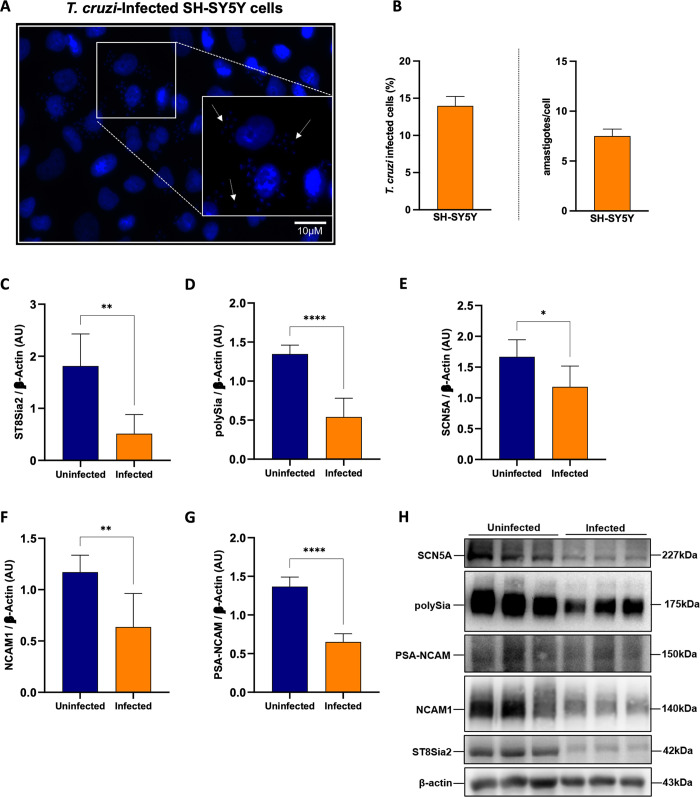
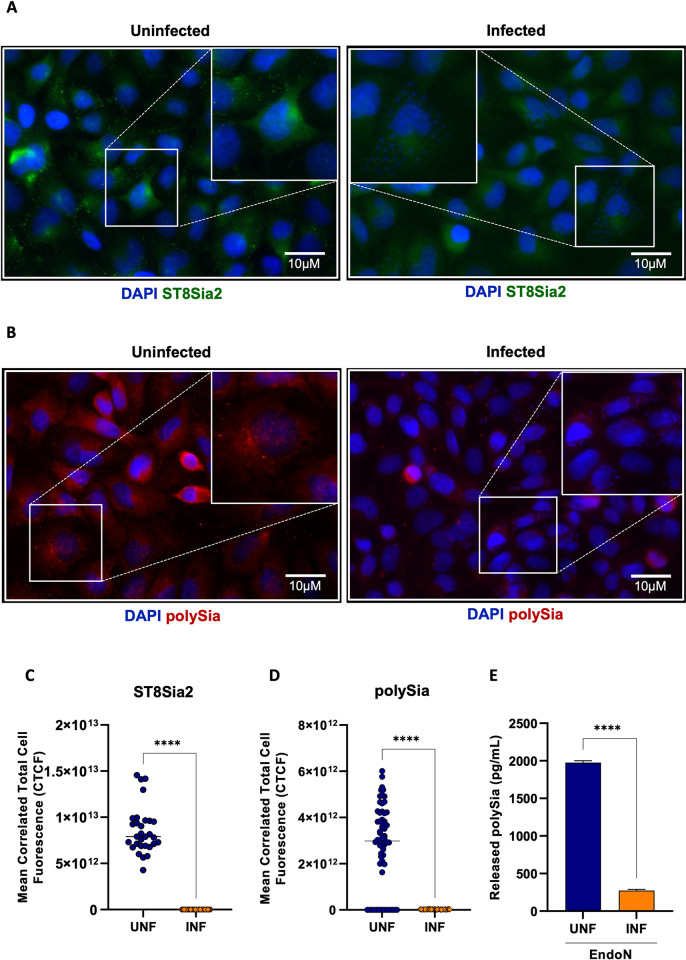
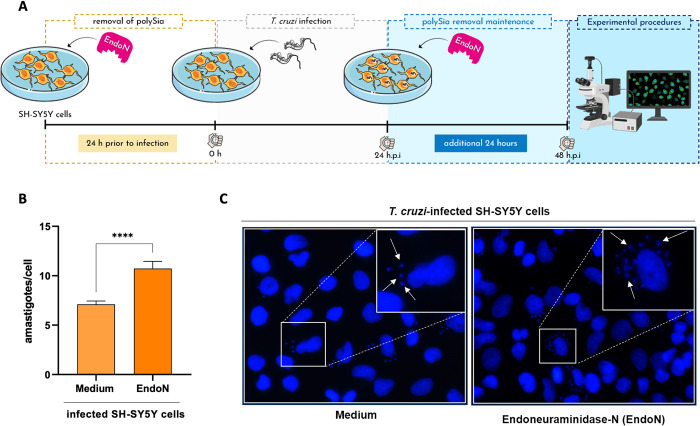
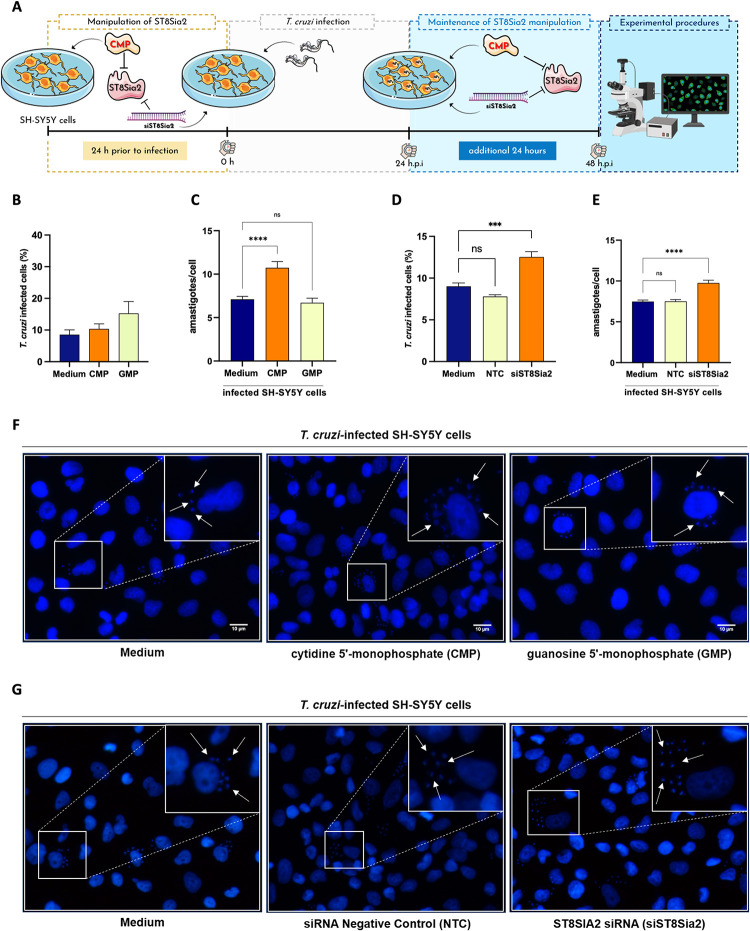
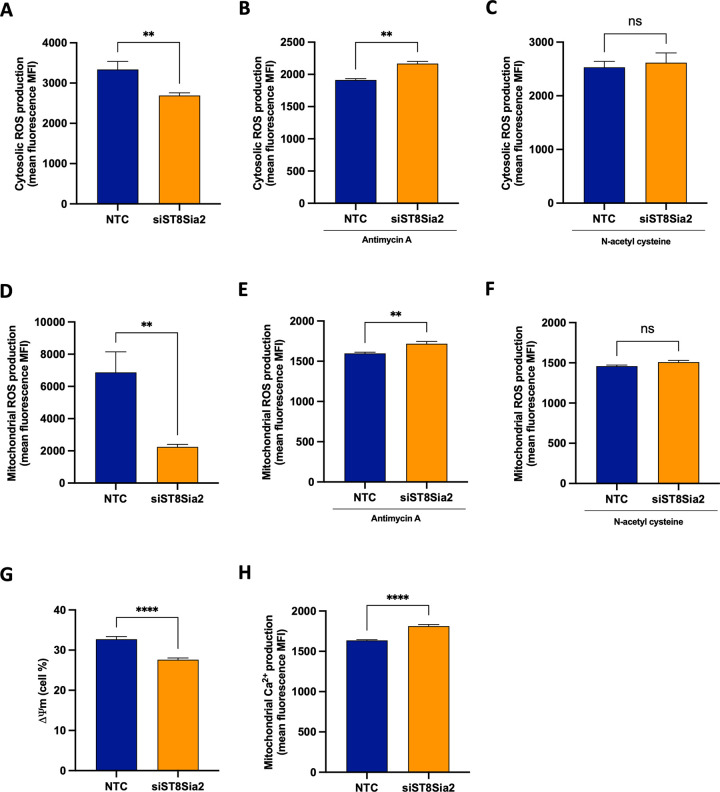
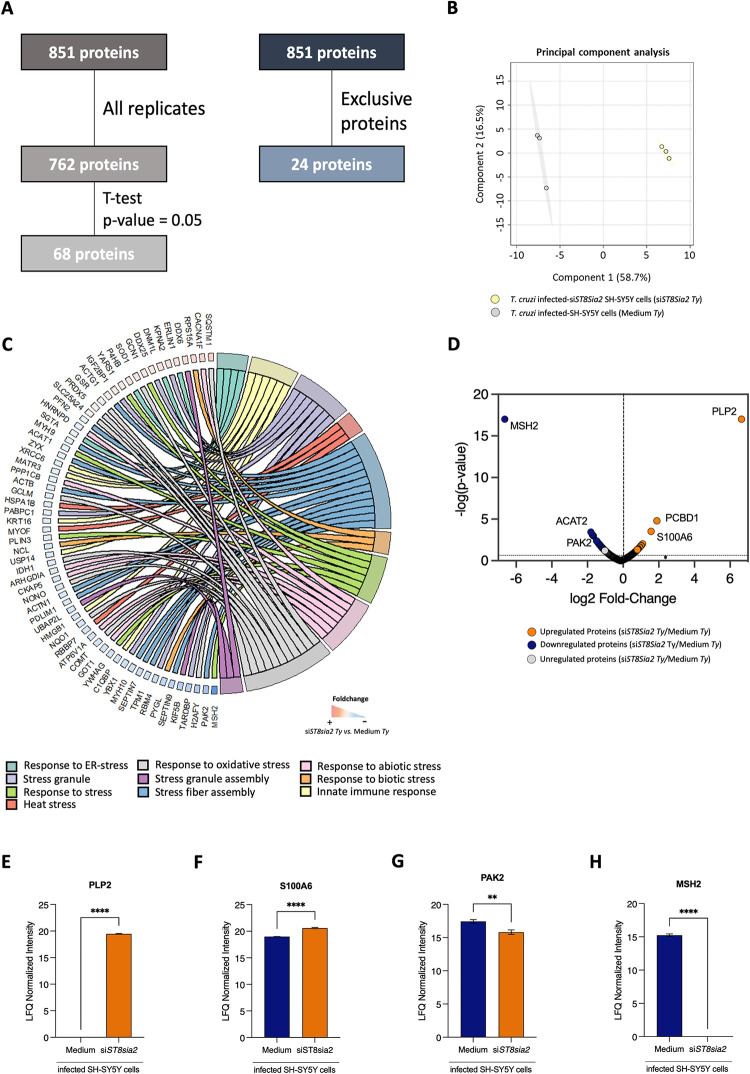
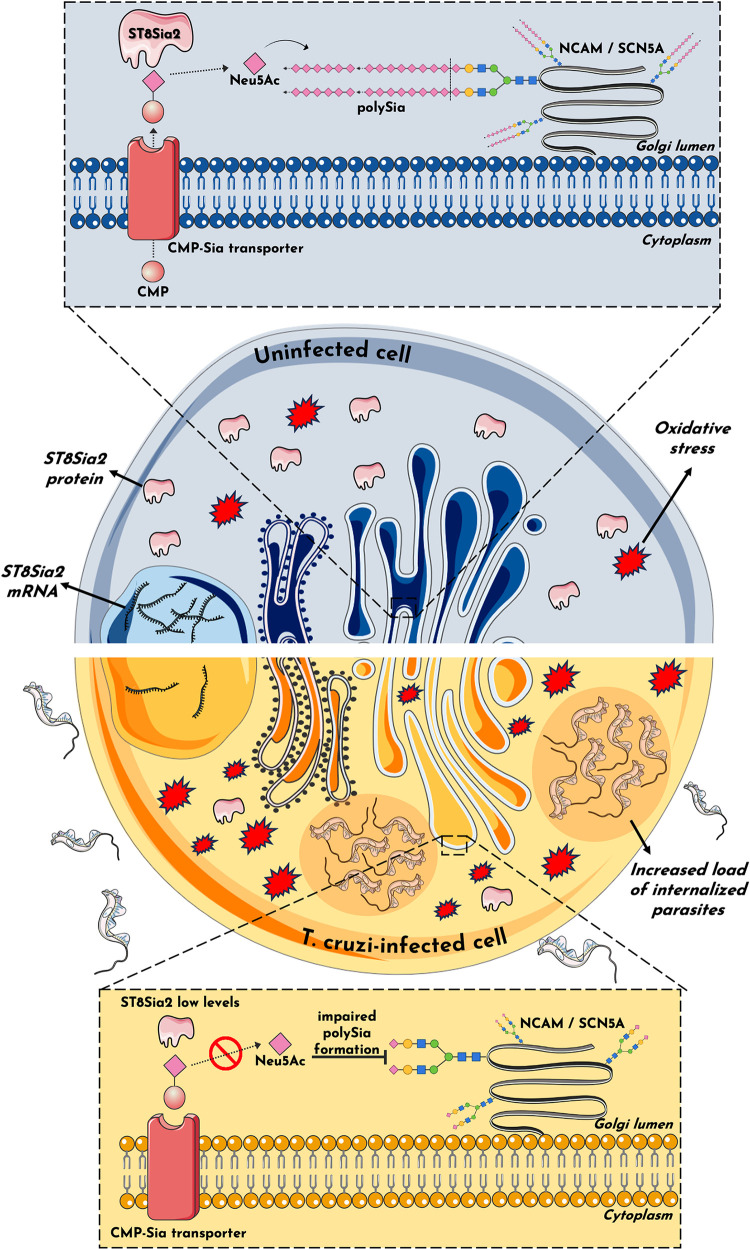
Glycosylation is one of the most structurally and functionally diverse co- and post-translational modifications in a cell. Addition and removal of glycans, especially to proteins and lipids, characterize this process which has important implications in several biological processes. In mammals, the repeated enzymatic addition of a sialic acid unit to underlying sialic acids (Sia) by polysialyltransferases, including ST8Sia2, leads to the formation of a sugar polymer called polysialic acid (polySia). The functional relevance of polySia has been extensively demonstrated in the nervous system. However, the role of polysialylation in infection is still poorly explored. Previous reports have shown that Trypanosoma cruzi (T. cruzi), a flagellated parasite that causes Chagas disease (CD), changes host sialylation of glycoproteins. To understand the role of host polySia during T. cruzi infection, we used a combination of in silico and experimental tools. We observed that T. cruzi reduces both the expression of the ST8Sia2 and the polysialylation of target substrates. We also found that chemical and genetic inhibition of host ST8Sia2 increased the parasite load in mammalian cells. We found that modulating host polysialylation may induce oxidative stress, creating a microenvironment that favors T. cruzi survival and infection. These findings suggest a novel approach to interfere with parasite infections through modulation of host polysialylation.
Copyright: © 2024 Barboza et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
