

Preclinical Evaluation of AZD6422, an Armored Chimeric Antigen Receptor T Cell Targeting CLDN18.2 in Gastric, Pancreatic, and Esophageal Cancers
- PMID: 39321207
- PMCID: PMC11609629
- DOI: 10.1158/1078-0432.CCR-24-1853
Preclinical Evaluation of AZD6422, an Armored Chimeric Antigen Receptor T Cell Targeting CLDN18.2 in Gastric, Pancreatic, and Esophageal Cancers
Abstract
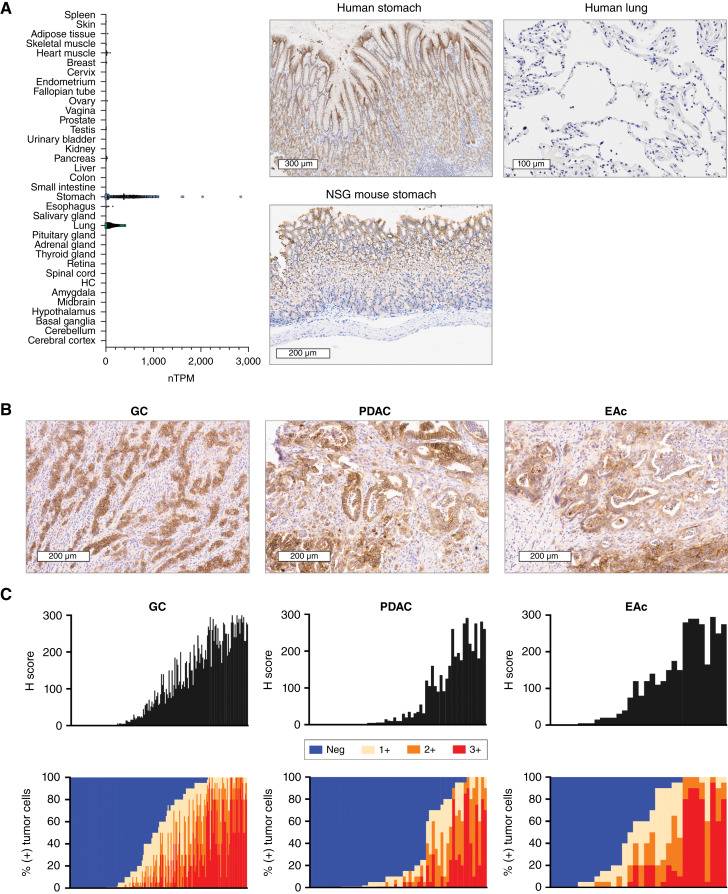
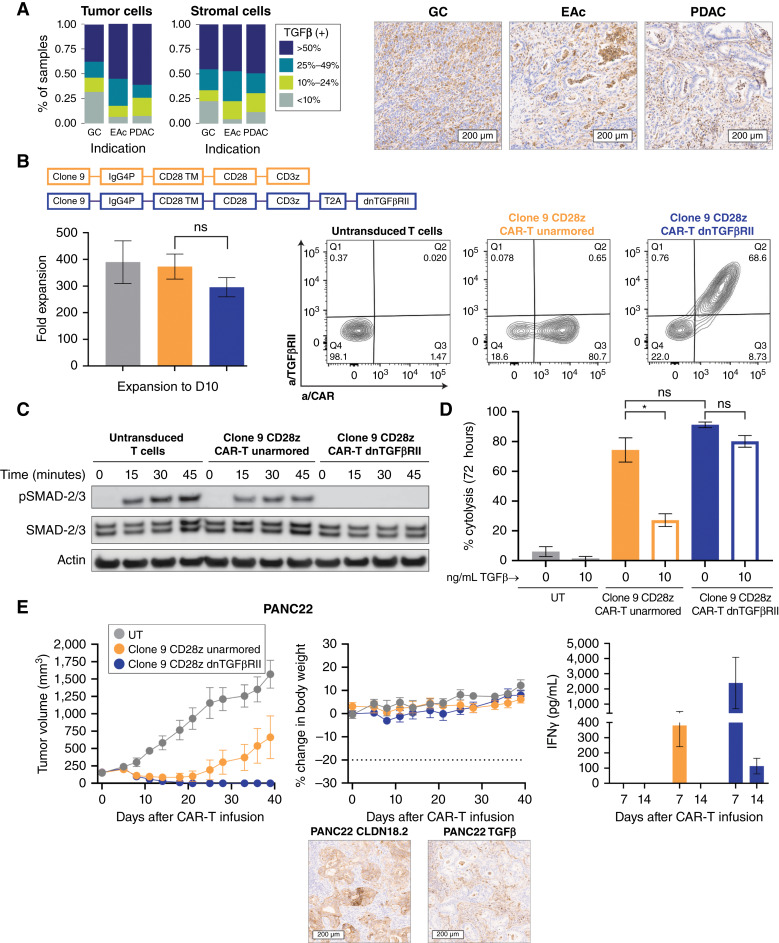
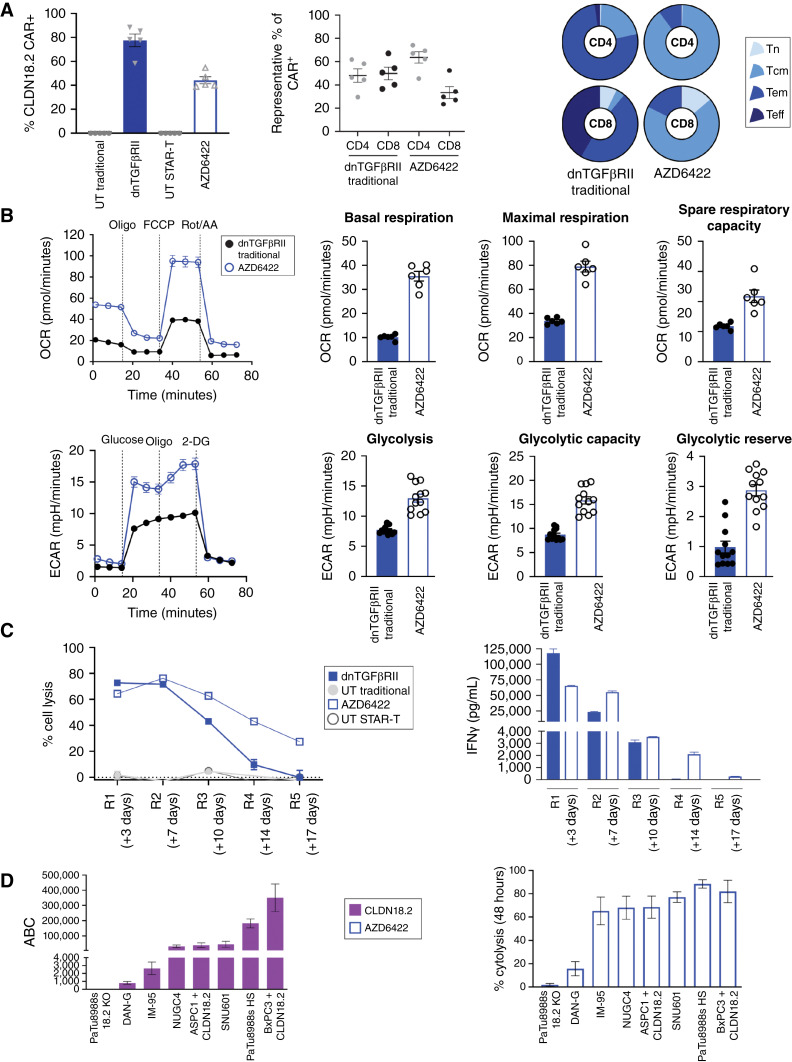
Purpose: Claudin 18.2 (CLDN18.2) is a surface membrane protein that is crucial for maintaining tight junctions in gastric mucosal cells and is highly expressed in gastric, esophageal, and pancreatic cancers. Thus, CLDN18.2 is suited for exploration as a clinical target for chimeric antigen receptor T-cell (CAR-T) therapy in these indications. Although CAR-T therapies show promise, a challenge faced in their development for solid tumors is the immunosuppressive tumor microenvironment, which is often characterized by the presence of immune and stromal cells secreting high levels of TGFβ. The addition of TGFβ armoring can potentially expand CAR-T activity in solid tumors. We report on the preclinical development of a CLDN18.2-targeting CAR-T therapy showing effectiveness in patient models with CLDN18.2-positive gastric, esophageal, and pancreatic tumors.
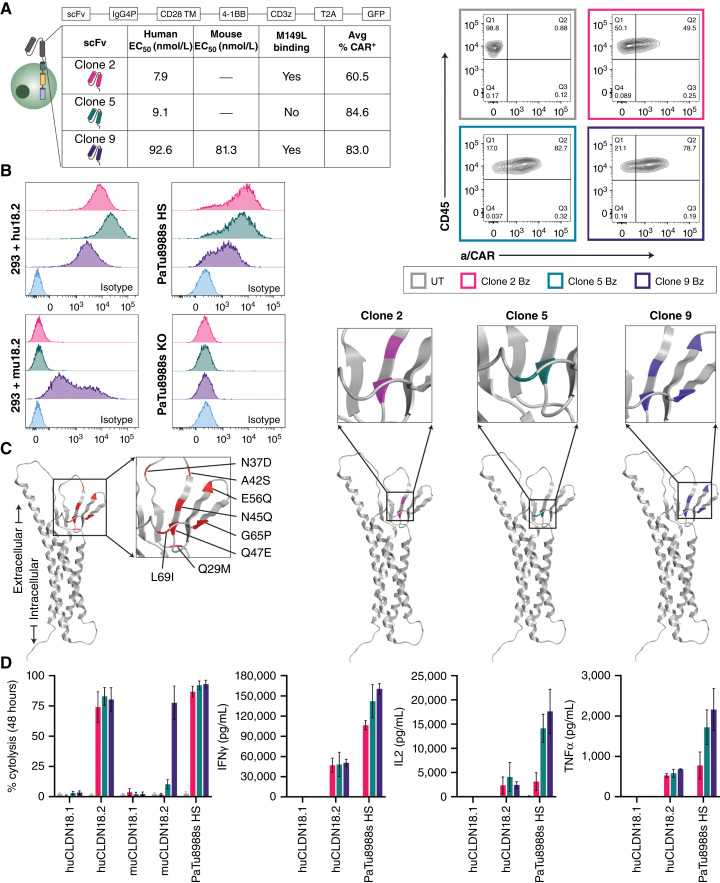
Experimental design: The lead lentivirus product contains a unique single-chain variable fragment; CD28 and CD3z costimulatory and signaling domains; and dominant-negative TGF-β receptor armoring, enhancing targeting and safety and counteracting suppression. We developed a shortened cell manufacturing process to enhance the potency of the final product AZD6422.
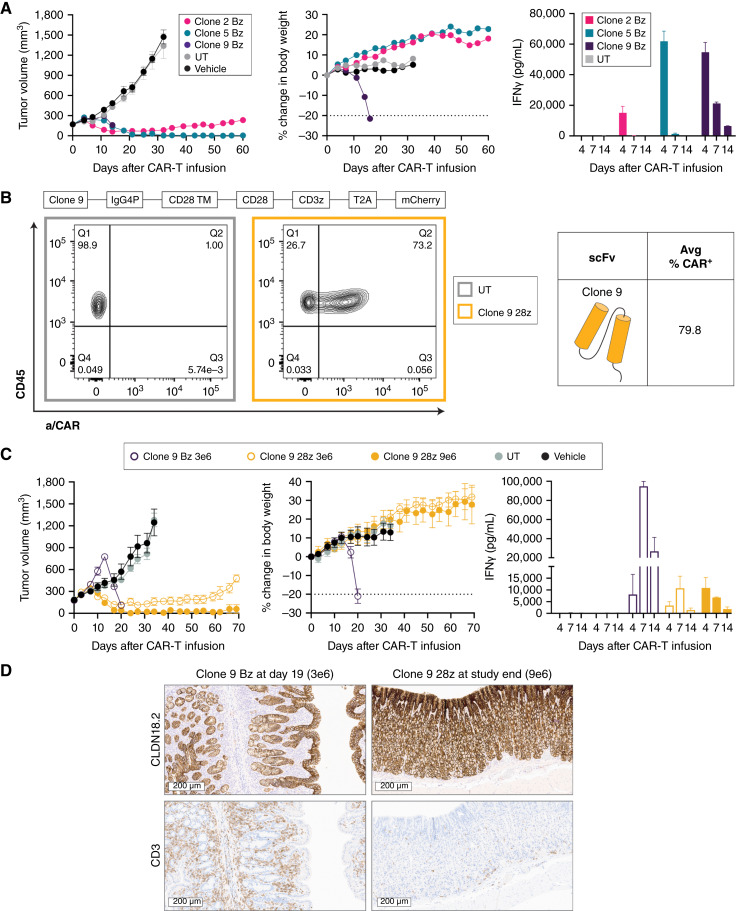
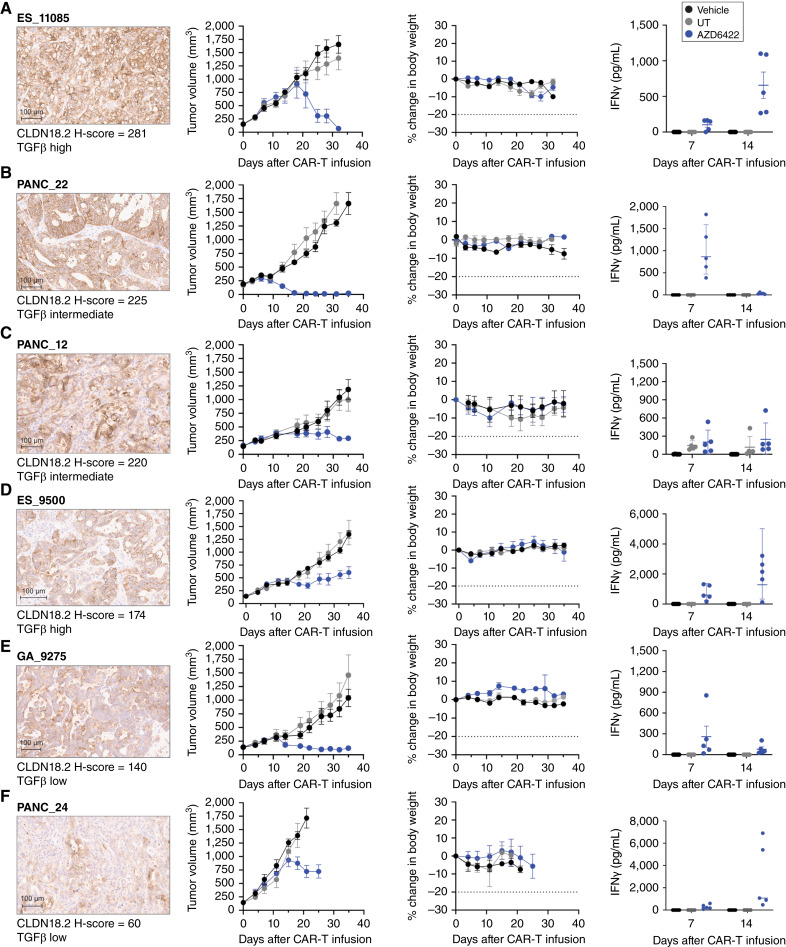
Results: AZD6422 exhibited significant antitumor activity and tolerability in multiple patient-derived tumor xenograft models with various CLDN18.2 and TGF-β levels, as determined by IHC. The efficacy of armored CAR-T cells in tumor models with elevated TGFβ was increased in vitro and in vivo. In vitro restimulation assays established greater persistence and cytolytic function of AZD6422 compared with a traditionally manufactured CAR-T.
Conclusions: AZD6422 was safe and efficacious in patient-derived, CLDN18.2-positive murine models of gastrointestinal cancers. Our data support further clinical development of AZD6422 for patients with these cancers.
©2024 The Authors; Published by the American Association for Cancer Research.
Conflict of interest statement
All authors were employees of AstraZeneca when the work was completed and may have stock ownership, interests, and/or options in the company. A.M. Barrett reports a patent for US20240228610A9 pending. Z.T. Britton reports other support from AstraZeneca outside the submitted work; a patent for US20240228610A9 pending; and employment with AstraZeneca/MedImmune. R.A. Carrasco reports a patent for US20240228610A9 pending. S. Breen reports a patent for US20240228610A9 pending. A.L. Hatke reports a patent for US20240228610A9 pending. C. Yang reports a patent for US20240228610A9 pending. P.L. Martin reports other support from AstraZeneca during the conduct of the study; other support from AstraZeneca outside the submitted work; and employment with AstraZeneca and ownership of AZ stock. J.-M. Lapointe reports personal fees from AstraZeneca during the conduct of the study. M. Cobbold reports other support from AstraZeneca during the conduct of the study. G. Moody reports a patent for US20240228610A9 pending to AstraZeneca, as well as employment with AstraZeneca LP and ownership of AstraZeneca stock. E.E. Bosco reports other support from AstraZeneca during the conduct of the study. No disclosures were reported by the other authors.
Figures
References
-
- Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. . Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin 2021;71:209–49. - PubMed
-
- Luo W, Wang J, Chen H, Qiu J, Wang R, Liu Y, et al. . Novel strategies optimize immunotherapy by improving the cytotoxic function of T cells for pancreatic cancer treatment. Cancer Lett 2023;576:216423. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
