

Offline hippocampal reactivation during dentate spikes supports flexible memory
- PMID: 39321790
- PMCID: PMC7616703
- DOI: 10.1016/j.neuron.2024.08.022
Offline hippocampal reactivation during dentate spikes supports flexible memory
Abstract
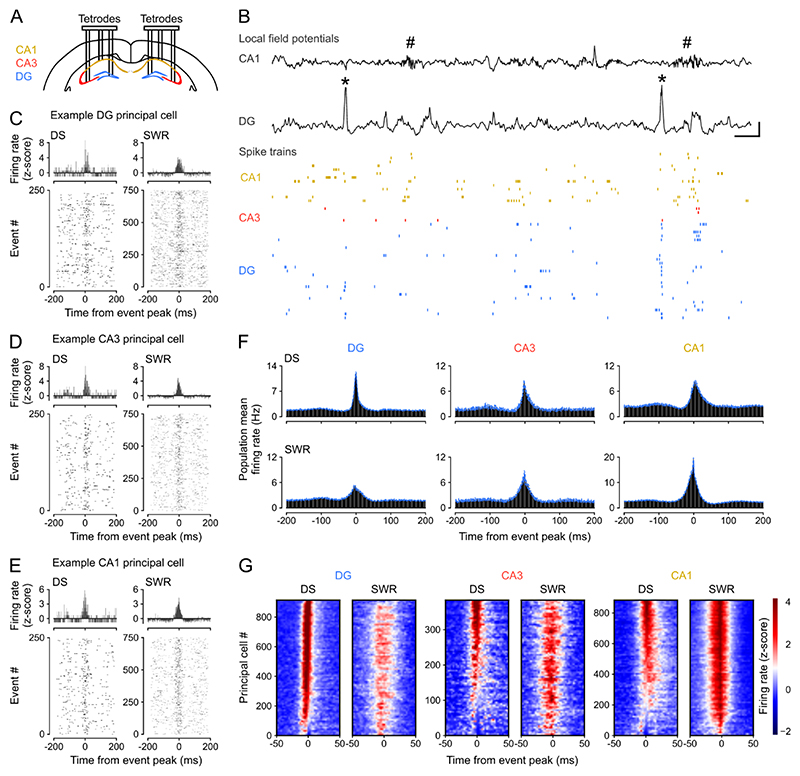
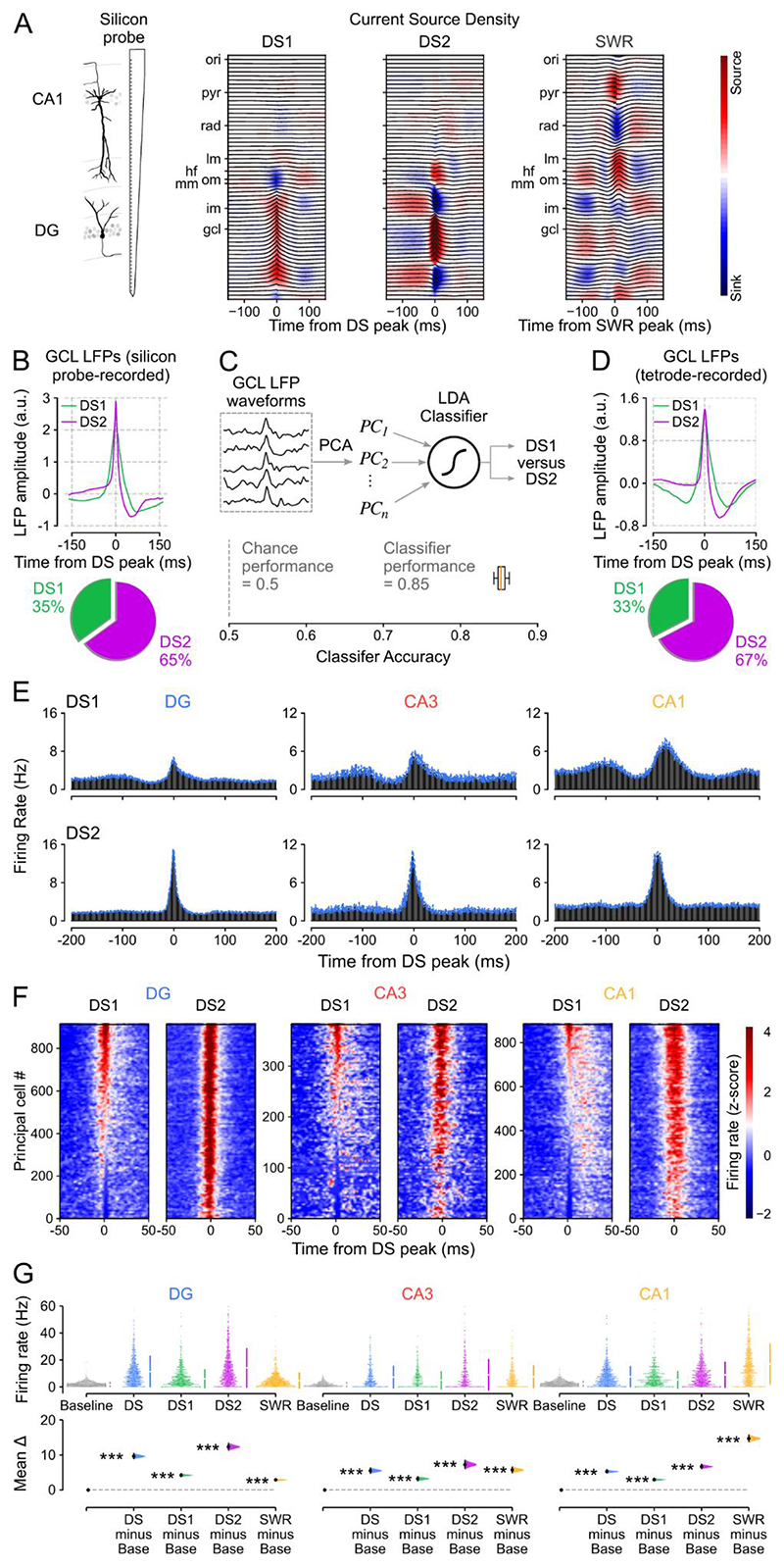
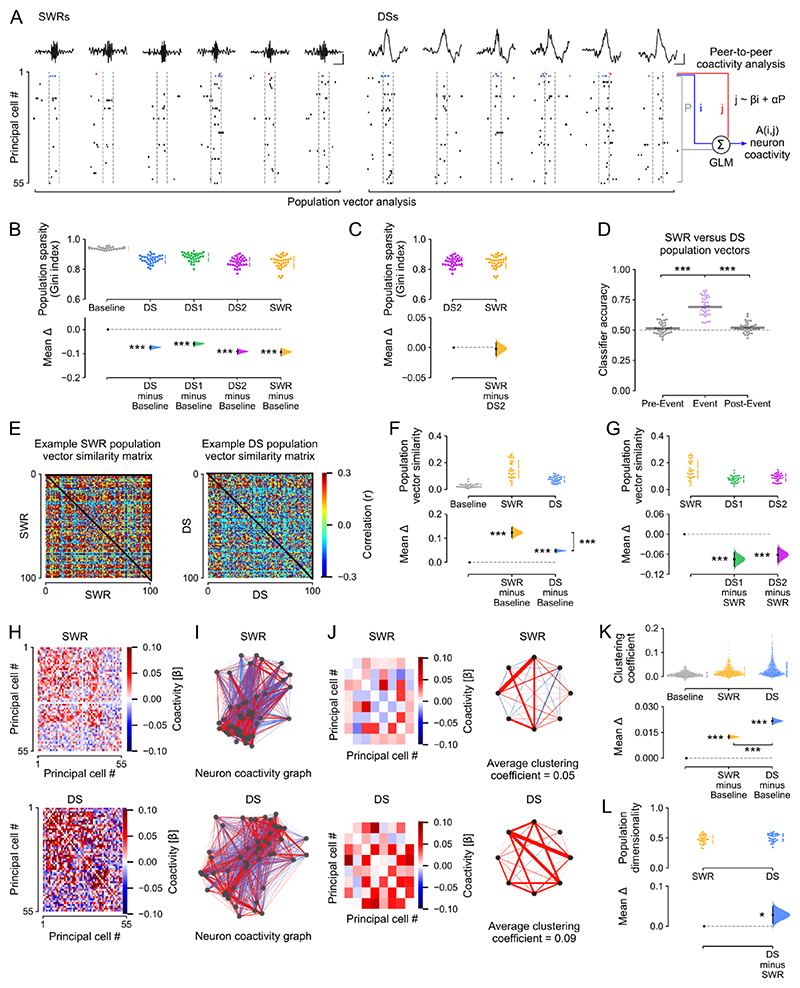
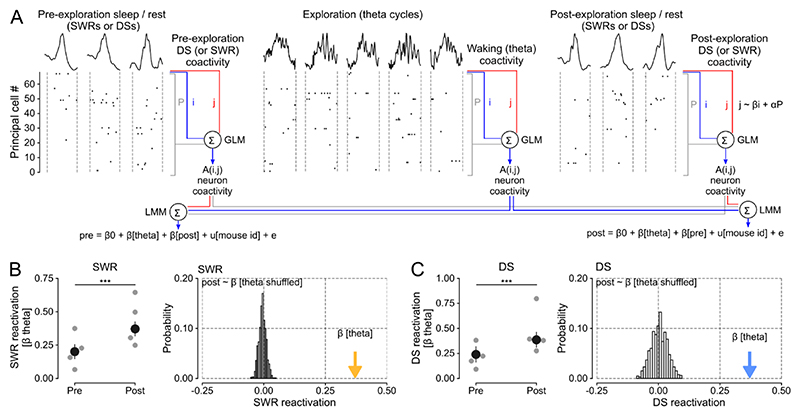
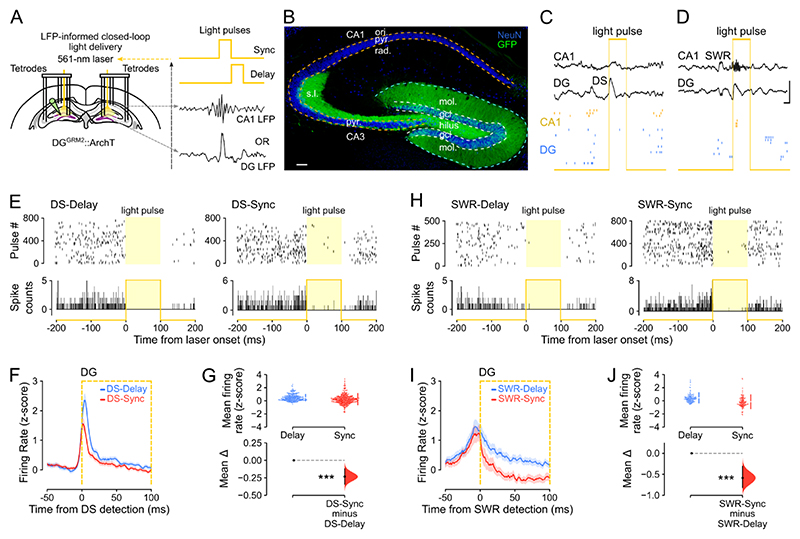
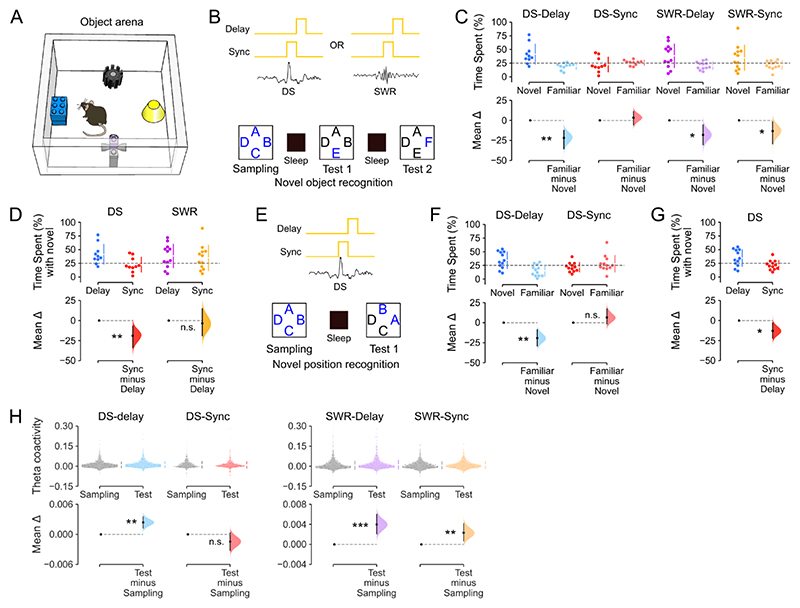
Stabilizing new memories requires coordinated neuronal spiking activity during sleep. Hippocampal sharp-wave ripples (SWRs) in the cornu ammonis (CA) region and dentate spikes (DSs) in the dentate gyrus (DG) are prime candidate network events for supporting this offline process. SWRs have been studied extensively, but the contribution of DSs remains unclear. By combining triple-ensemble (DG-CA3-CA1) recordings and closed-loop optogenetics in mice, we show that, like SWRs, DSs synchronize spiking across DG and CA principal cells to reactivate population-level patterns of neuronal coactivity expressed during prior waking experience. Notably, the population coactivity structure in DSs is more diverse and higher dimensional than that seen during SWRs. Importantly, suppressing DG granule cell spiking selectively during DSs impairs subsequent flexible memory performance during multi-object recognition tasks and associated hippocampal patterns of neuronal coactivity. We conclude that DSs constitute a second offline network event central to hippocampal population dynamics serving memory-guided behavior.
Keywords: dentate spikes; hippocampus; memory consolidation; neuronal coactivity; offline reactivation; population patterns; sharp-wave ripples.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Maquet P. The Role of Sleep in Learning and Memory. Science. 2001;294:1048–1052. - PubMed
-
- Walker MP, Stickgold R. Sleep, Memory, and Plasticity. Annual Review of Psychology. 2006;57:139–166. - PubMed
-
- Klinzing JG, Niethard N, Born J. Mechanisms of systems memory consolidation during sleep. Nat Neurosci. 2019;22:1598–1610. - PubMed
-
- Brodt S, Inostroza M, Niethard N, Born J. Sleep—A brain-state serving systems memory consolidation. Neuron. 2023;111:1050–1075. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
