

Engineered allogeneic T cells decoupling T-cell-receptor and CD3 signalling enhance the antitumour activity of bispecific antibodies
- PMID: 39322719
- PMCID: PMC11668682
- DOI: 10.1038/s41551-024-01255-x
Engineered allogeneic T cells decoupling T-cell-receptor and CD3 signalling enhance the antitumour activity of bispecific antibodies
Abstract
Bispecific antibodies (biAbs) used in cancer immunotherapies rely on functional autologous T cells, which are often damaged and depleted in patients with haematological malignancies and in other immunocompromised patients. The adoptive transfer of allogeneic T cells from healthy donors can enhance the efficacy of biAbs, but donor T cells binding to host-cell antigens cause an unwanted alloreactive response. Here we show that allogeneic T cells engineered with a T-cell receptor that does not convert antigen binding into cluster of differentiation 3 (CD3) signalling decouples antigen-mediated T-cell activation from T-cell cytotoxicity while preserving the surface expression of the T-cell-receptor-CD3 signalling complex as well as biAb-mediated CD3 signalling and T-cell activation. In mice with CD19+ tumour xenografts, treatment with the engineered human cells in combination with blinatumomab (a clinically approved biAb) led to the recognition and clearance of tumour cells in the absence of detectable alloreactivity. Our findings support the development of immunotherapies combining biAbs and 'off-the-shelf' allogeneic T cells.
© 2024. The Author(s).
Conflict of interest statement
Competing interests: E.K., S.T.R., C.R.W. and R.V.-L. are co -inventors on the patent ‘Universal TCR variants for allogeneic immunotherapy’ filed by ETH Zurich related to AED TCRs and methods of their use. The other authors declare no competing interests.
Figures

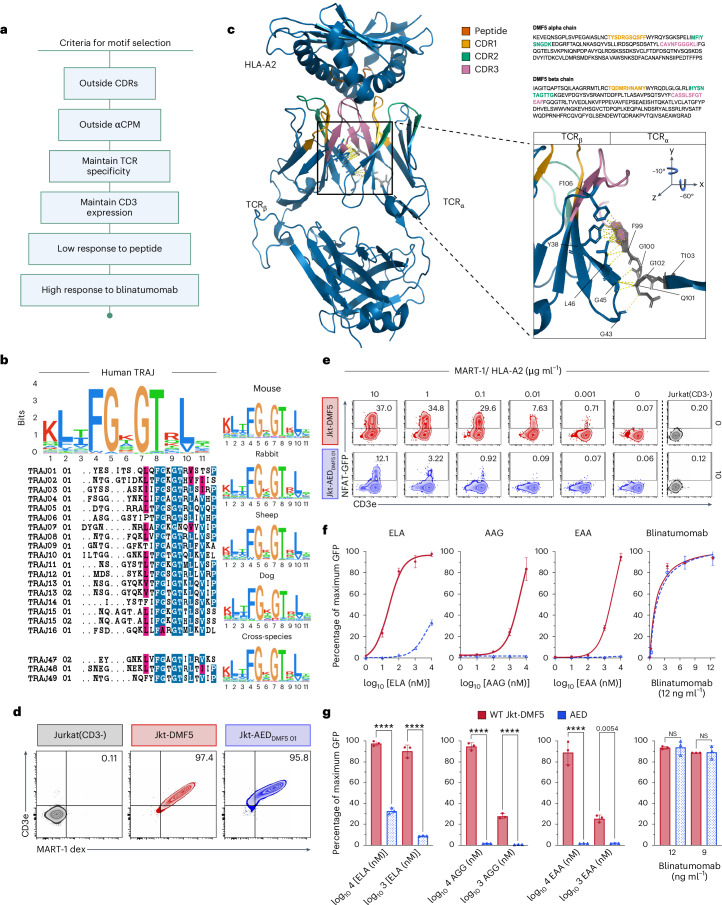

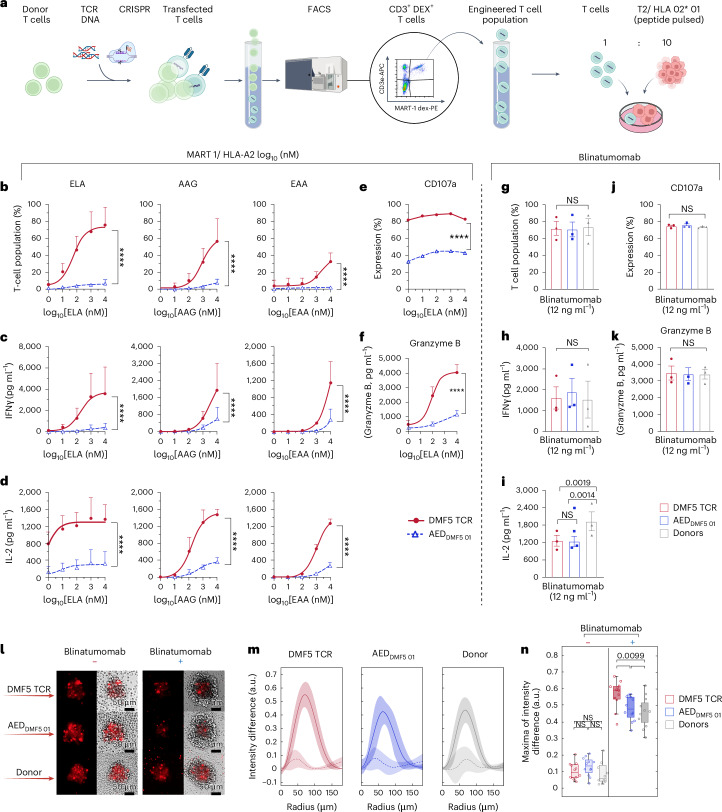
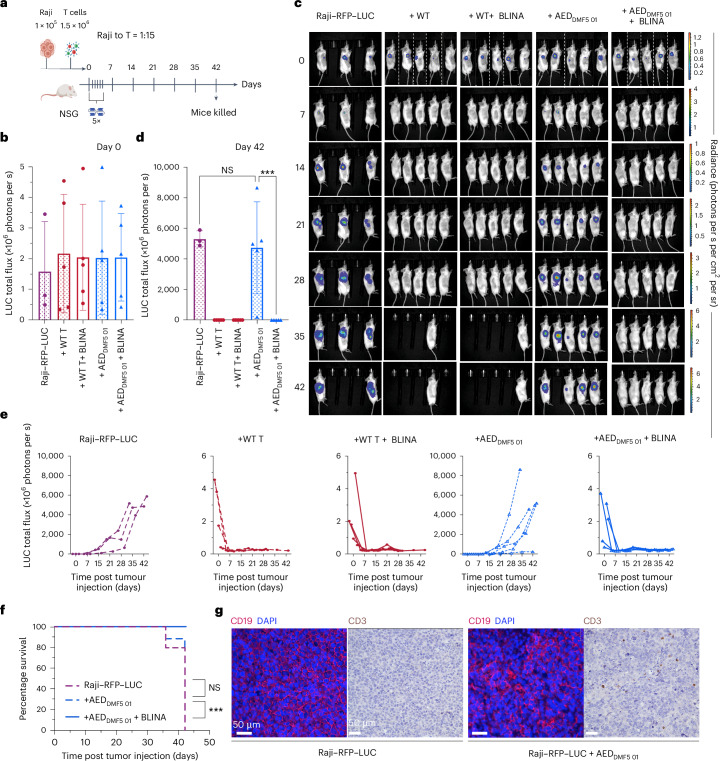
References
-
- Topp, M. S. et al. Phase II trial of the anti-CD19 bispecific T cell–engager blinatumomab shows hematologic and molecular remissions in patients with relapsed or refractory B-precursor acute lymphoblastic leukemia. J. Clin. Oncol.32, 4134–4140 (2014). - PubMed
-
- Jabbour, E. et al. Outcome of patients with relapsed/refractory acute lymphoblastic leukemia after blinatumomab failure: no change in the level of CD19 expression. Am. J. Hematol.93, 371–374 (2018). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
