

Neuromodulator and neuropeptide sensors and probes for precise circuit interrogation in vivo
- PMID: 39325905
- PMCID: PMC11488521
- DOI: 10.1126/science.adn6671
Neuromodulator and neuropeptide sensors and probes for precise circuit interrogation in vivo
Abstract
To determine how neuronal circuits encode and drive behavior, it is often necessary to measure and manipulate different aspects of neurochemical signaling in awake animals. Optogenetics and calcium sensors have paved the way for these types of studies, allowing for the perturbation and readout of spiking activity within genetically defined cell types. However, these methods lack the ability to further disentangle the roles of individual neuromodulator and neuropeptides on circuits and behavior. We review recent advances in chemical biology tools that enable precise spatiotemporal monitoring and control over individual neuroeffectors and their receptors in vivo. We also highlight discoveries enabled by such tools, revealing how these molecules signal across different timescales to drive learning, orchestrate behavioral changes, and modulate circuit activity.
Conflict of interest statement
Figures
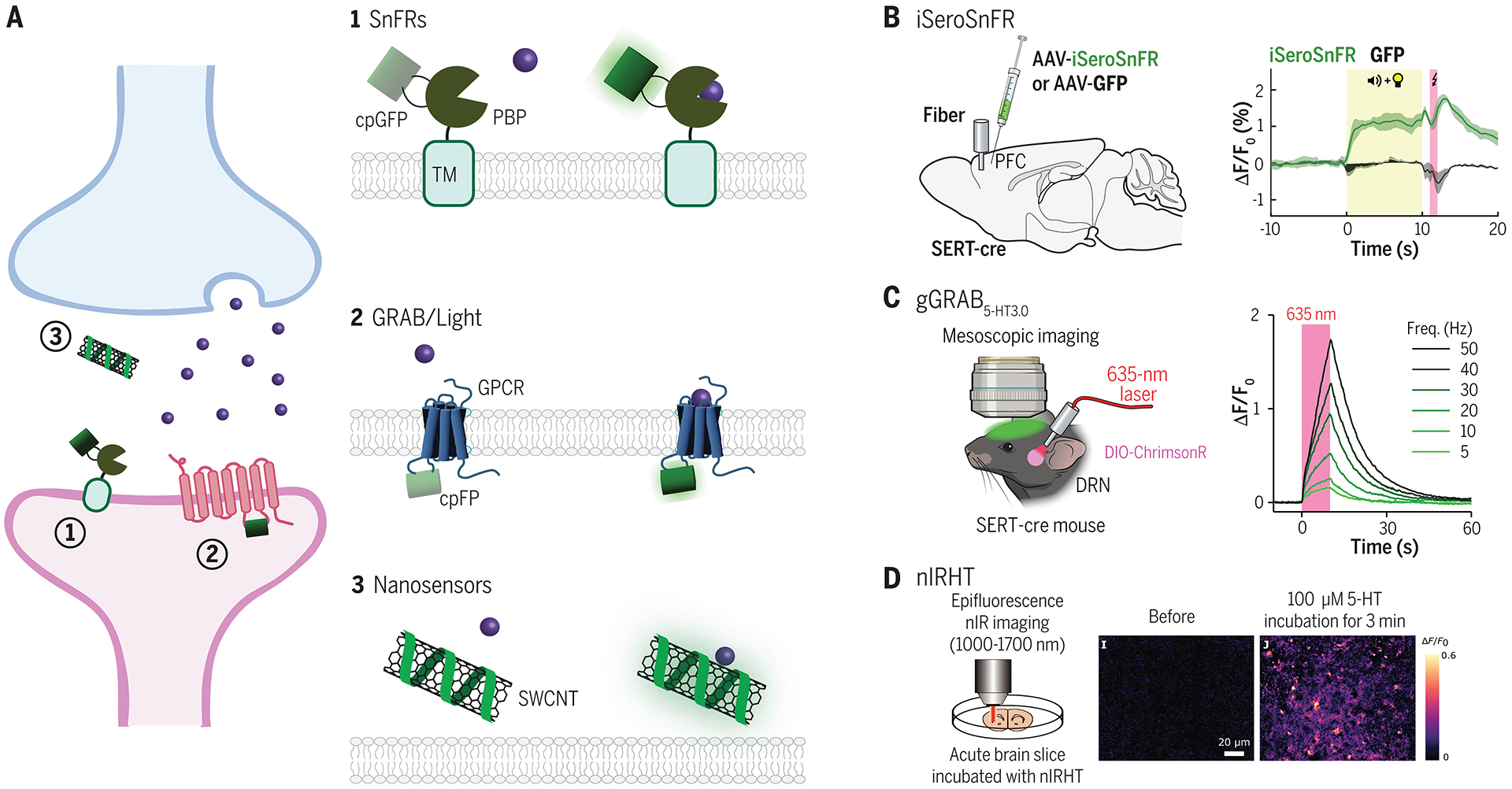
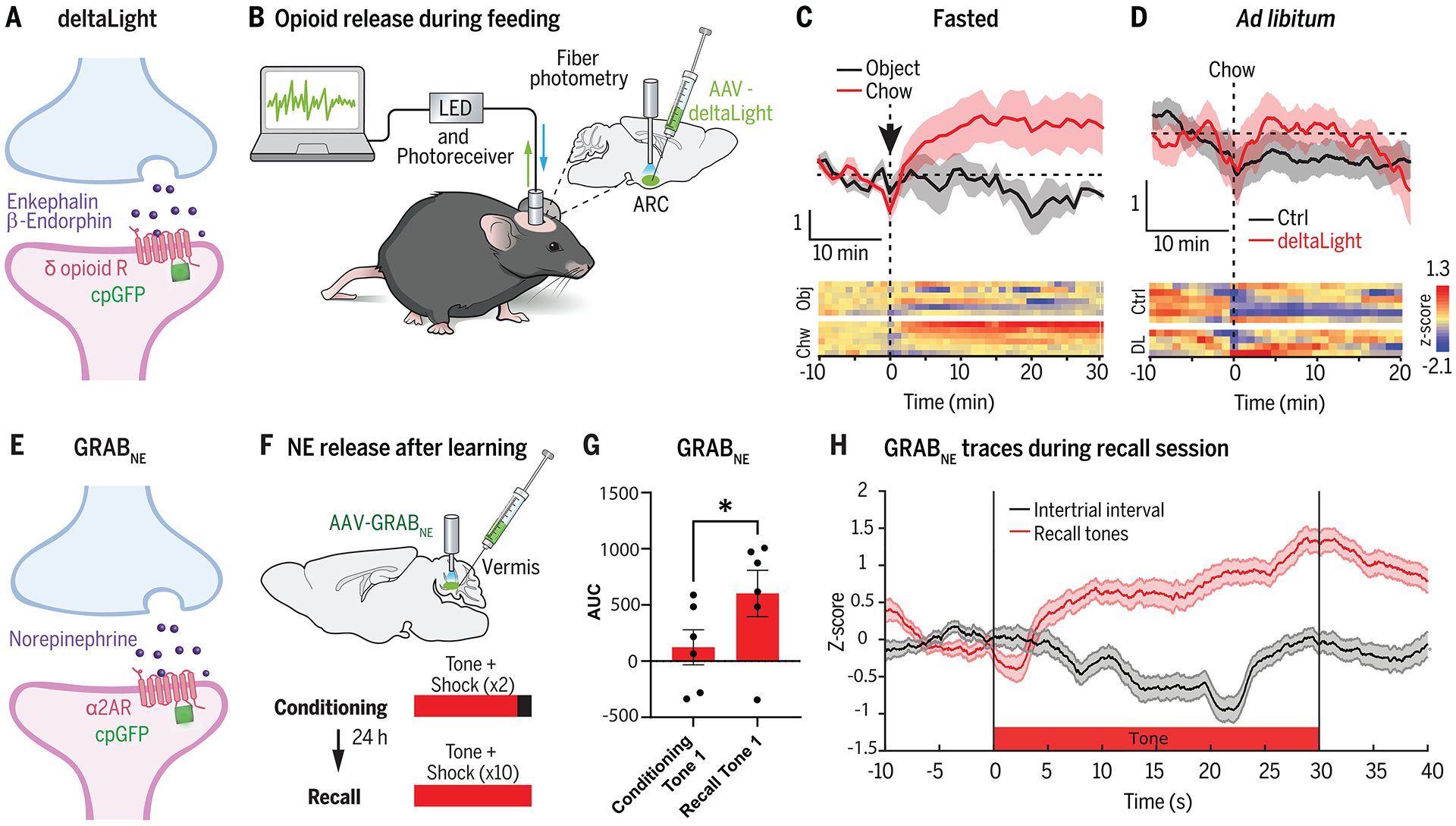
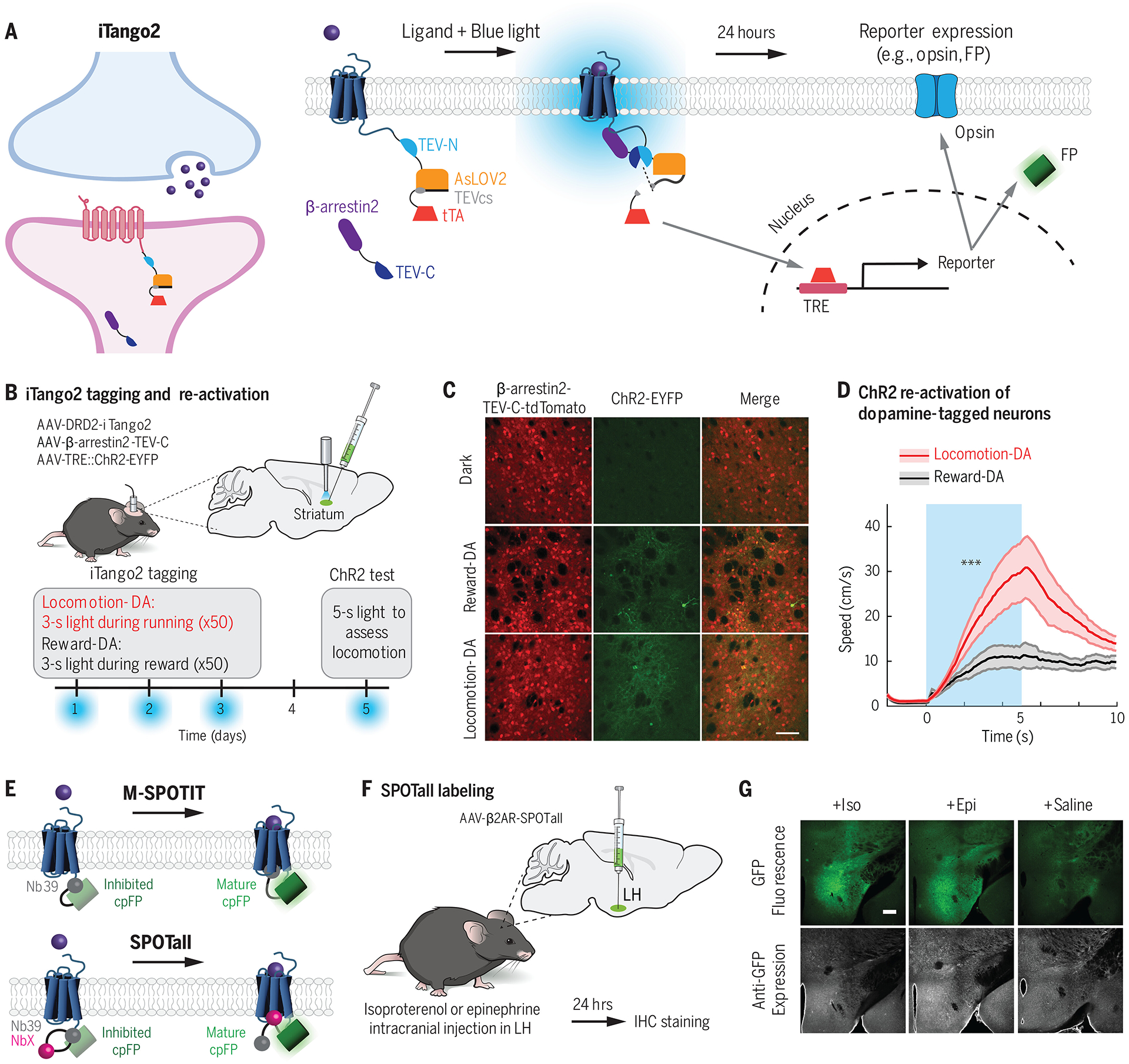
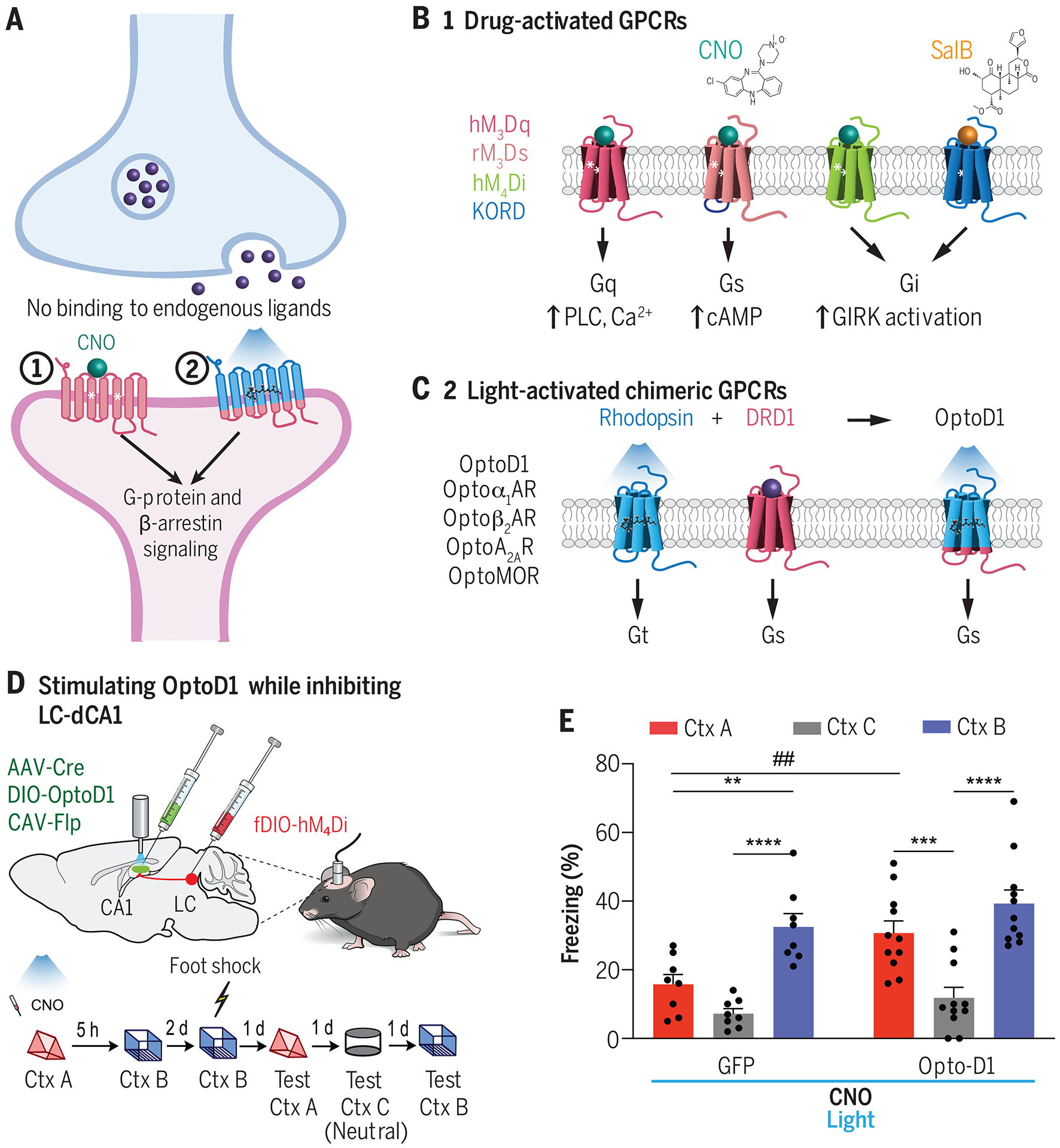
Similar articles
-
Ultrafast neuronal imaging of dopamine dynamics with designed genetically encoded sensors.Science. 2018 Jun 29;360(6396):eaat4422. doi: 10.1126/science.aat4422. Epub 2018 May 31. Science. 2018. PMID: 29853555 Free PMC article.
-
Imaging Neurotransmitter and Neuromodulator Dynamics In Vivo with Genetically Encoded Indicators.Neuron. 2020 Oct 14;108(1):17-32. doi: 10.1016/j.neuron.2020.09.036. Neuron. 2020. PMID: 33058762 Review.
-
All-Optical Interrogation of Neural Circuits.J Neurosci. 2015 Oct 14;35(41):13917-26. doi: 10.1523/JNEUROSCI.2916-15.2015. J Neurosci. 2015. PMID: 26468193 Free PMC article.
-
A genetically encoded tool for reconstituting synthetic modulatory neurotransmission and reconnect neural circuits in vivo.Nat Commun. 2021 Aug 9;12(1):4795. doi: 10.1038/s41467-021-24690-9. Nat Commun. 2021. PMID: 34373460 Free PMC article.
-
Imaging voltage and brain chemistry with genetically encoded sensors and modulators.Curr Opin Chem Biol. 2020 Aug;57:166-176. doi: 10.1016/j.cbpa.2020.07.006. Epub 2020 Aug 18. Curr Opin Chem Biol. 2020. PMID: 32823064 Review.
Cited by
-
Illuminating the impact of stress: In vivo approaches to track stress-related neural adaptations.Neurobiol Stress. 2025 Feb 7;35:100712. doi: 10.1016/j.ynstr.2025.100712. eCollection 2025 Mar. Neurobiol Stress. 2025. PMID: 40191171 Free PMC article.
-
Principles and Design of Molecular Tools for Sensing and Perturbing Cell Surface Receptor Activity.Chem Rev. 2025 Mar 12;125(5):2665-2702. doi: 10.1021/acs.chemrev.4c00582. Epub 2025 Feb 25. Chem Rev. 2025. PMID: 39999110 Review.
-
The brain-heart axis: integrative cooperation of neural, mechanical and biochemical pathways.Nat Rev Cardiol. 2025 Aug;22(8):537-550. doi: 10.1038/s41569-025-01140-3. Epub 2025 Mar 3. Nat Rev Cardiol. 2025. PMID: 40033035 Review.
-
Coding principles and mechanisms of serotonergic transmission modes.Mol Psychiatry. 2025 Aug;30(8):3430-3442. doi: 10.1038/s41380-025-02930-4. Epub 2025 Feb 22. Mol Psychiatry. 2025. PMID: 39987232 Free PMC article.
-
Transient pre-seizure inhibition of lateral hypothalamic orexin neurons: a novel possibility for seizure control.MedComm (2020). 2025 Jan 14;6(2):e70070. doi: 10.1002/mco2.70070. eCollection 2025 Feb. MedComm (2020). 2025. PMID: 39822759 Free PMC article. No abstract available.
