

Structural genomic variation and behavioral interactions underpin a balanced sexual mimicry polymorphism
- PMID: 39326413
- PMCID: PMC12129486
- DOI: 10.1016/j.cub.2024.08.053
Structural genomic variation and behavioral interactions underpin a balanced sexual mimicry polymorphism
Abstract
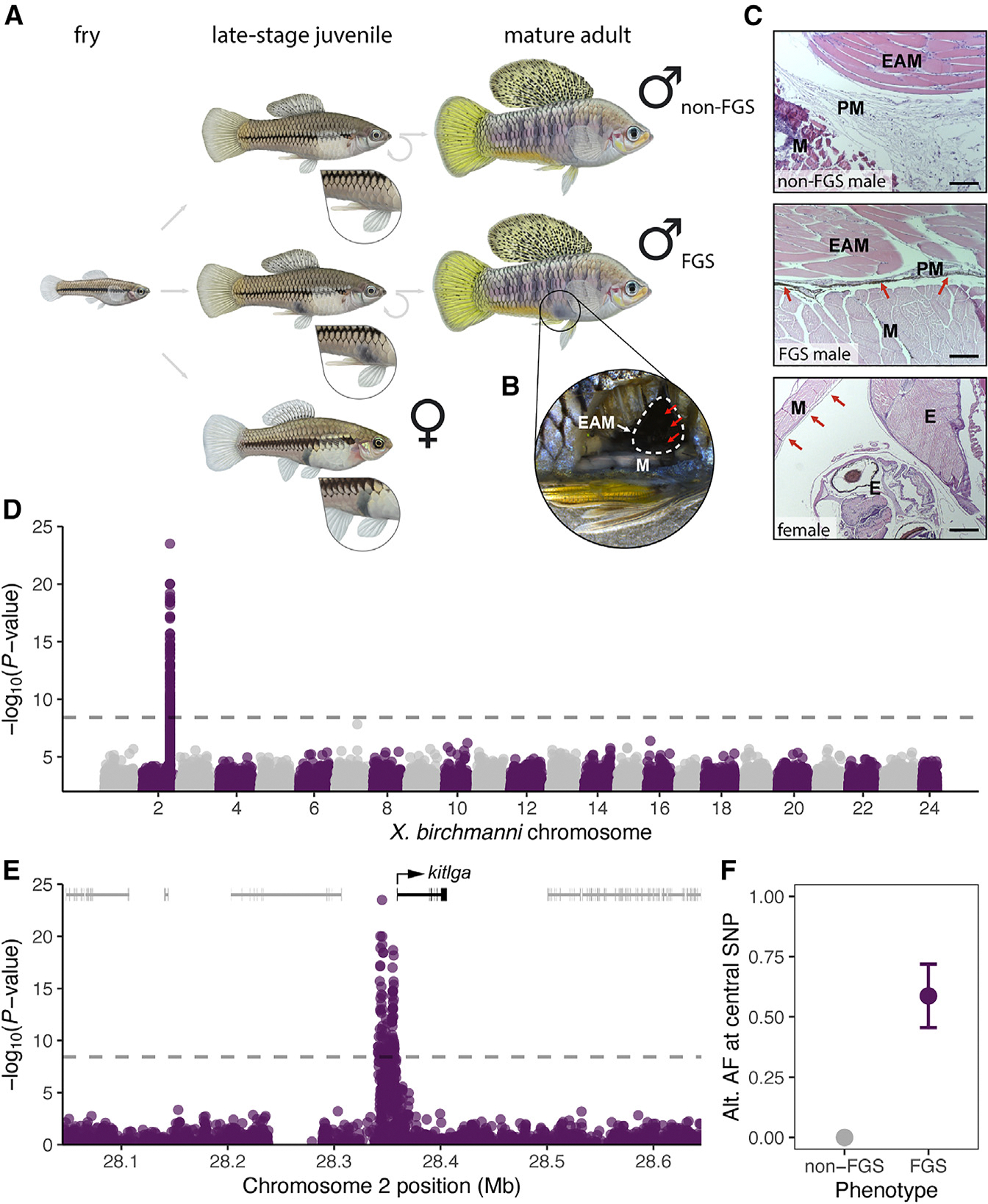
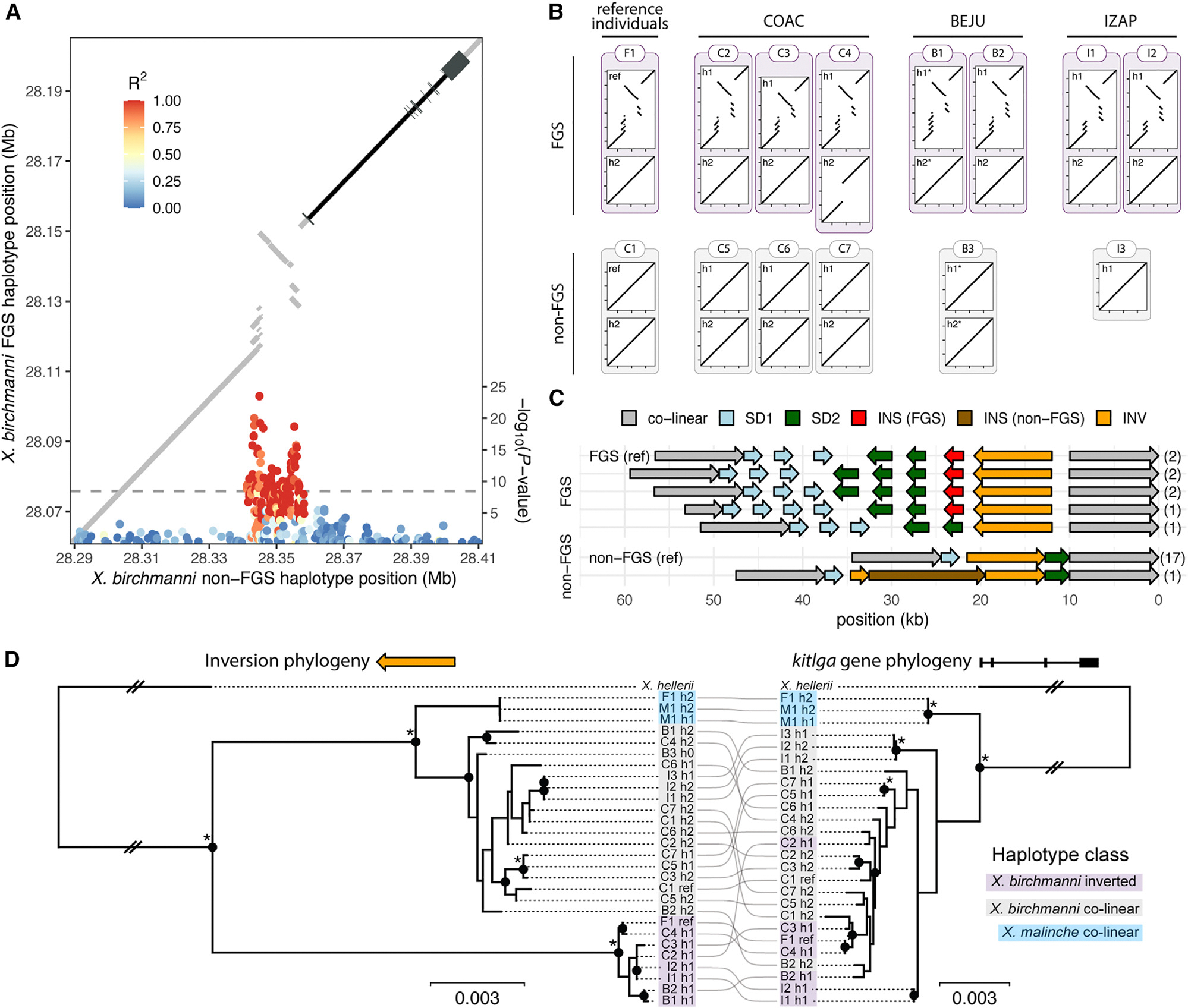
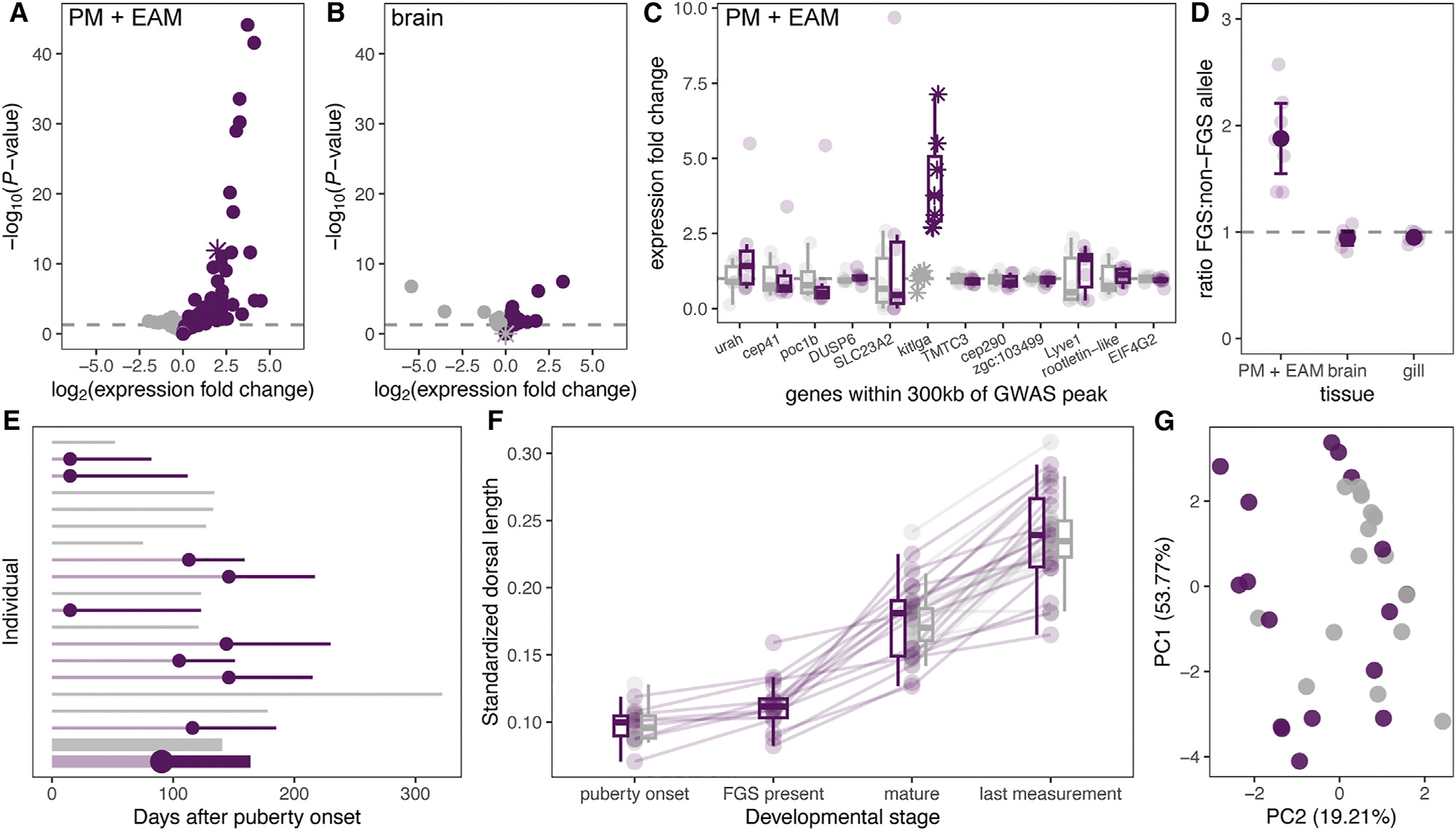
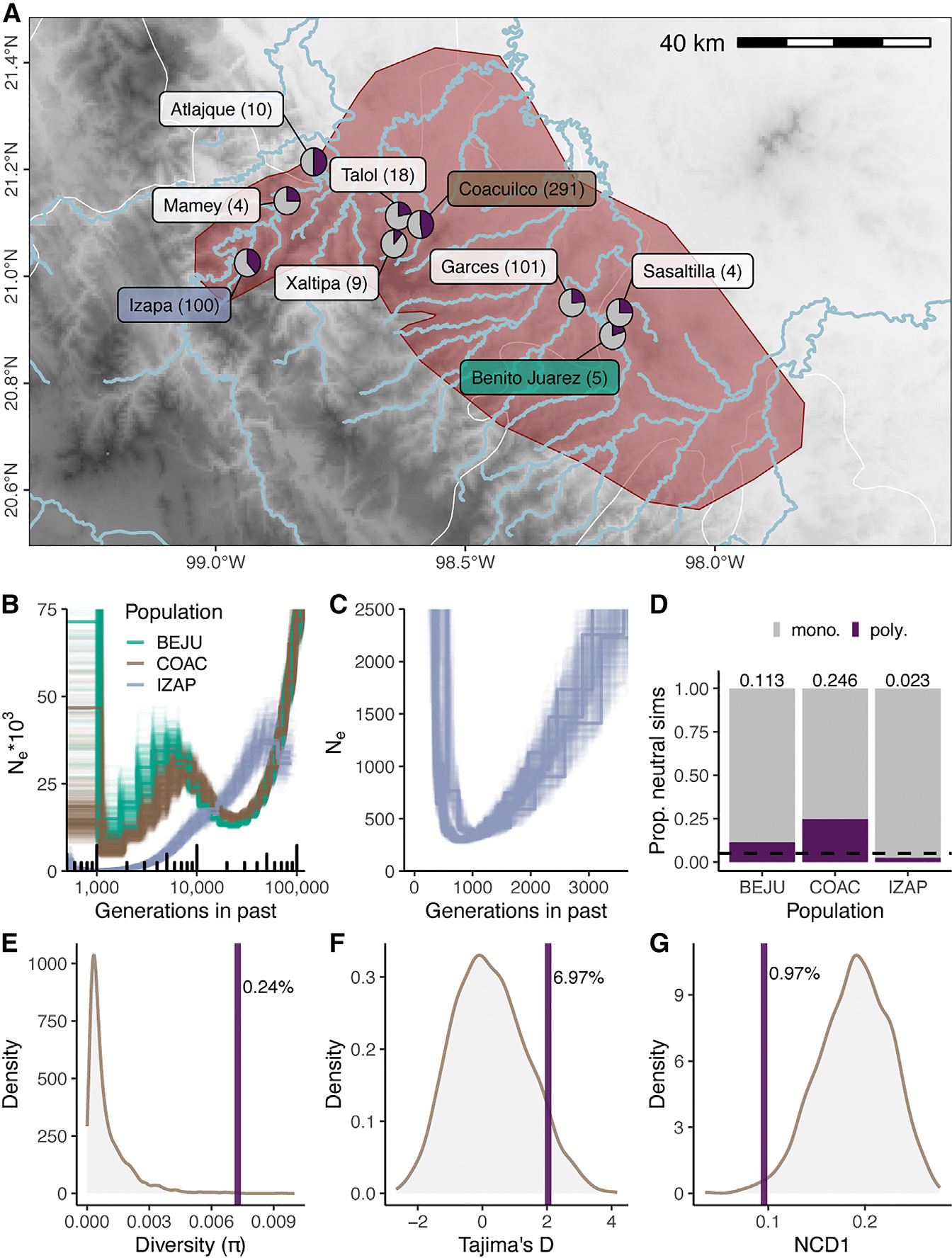
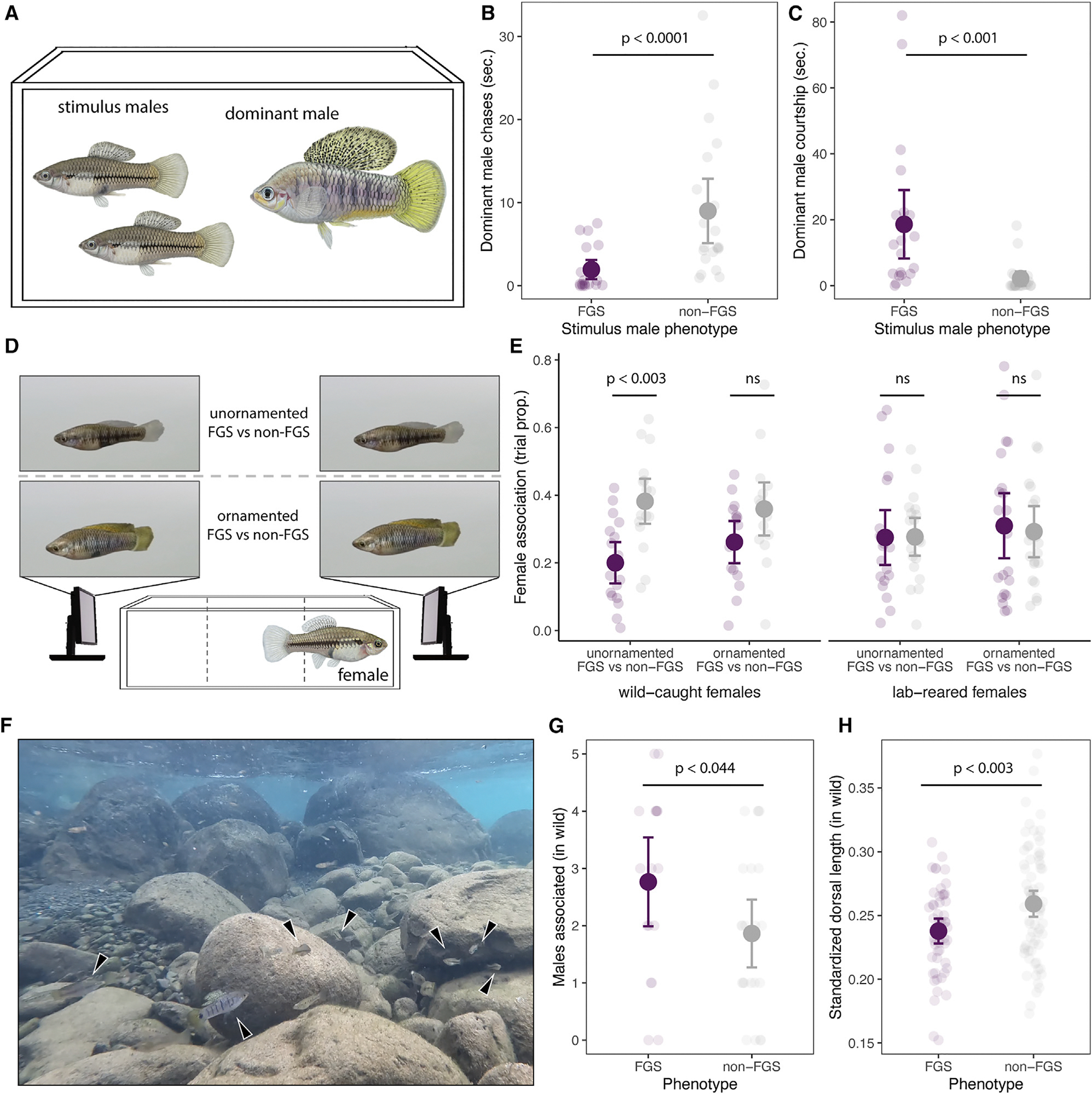
How phenotypic diversity originates and persists within populations are classic puzzles in evolutionary biology. While balanced polymorphisms segregate within many species, it remains rare for both the genetic basis and the selective forces to be known, leading to an incomplete understanding of many classes of traits under balancing selection. Here, we uncover the genetic architecture of a balanced sexual mimicry polymorphism and identify behavioral mechanisms that may be involved in its maintenance in the swordtail fish Xiphophorus birchmanni. We find that ∼40% of X. birchmanni males develop a "false gravid spot," a melanic pigmentation pattern that mimics the "pregnancy spot" associated with sexual maturity in female live-bearing fish. Using genome-wide association mapping, we detect a single intergenic region associated with variation in the false gravid spot phenotype, which is upstream of kitlga, a melanophore patterning gene. By performing long-read sequencing within and across populations, we identify complex structural rearrangements between alternate alleles at this locus. The false gravid spot haplotype drives increased allele-specific expression of kitlga, which provides a mechanistic explanation for the increased melanophore abundance that causes the spot. By studying social interactions in the laboratory and in nature, we find that males with the false gravid spot experience less aggression; however, they also receive increased attention from other males and are disdained by females. These behavioral interactions may contribute to the maintenance of this phenotypic polymorphism in natural populations. We speculate that structural variants affecting gene regulation may be an underappreciated driver of balanced polymorphisms across diverse species.
Keywords: Xiphophorus; balancing selection; kit ligand; polymorphism; sexual mimicry; structural variation.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Multiple mating and reproductive skew in parental and introgressed females of the live-bearing fish Xiphophorus birchmanni.J Hered. 2015 Jan-Feb;106(1):57-66. doi: 10.1093/jhered/esu066. Epub 2014 Nov 27. J Hered. 2015. PMID: 25433083 Free PMC article.
-
Sex-specific plasticity and genotype × sex interactions for age and size of maturity in the sheepshead swordtail, Xiphophorus birchmanni.J Evol Biol. 2016 Mar;29(3):645-56. doi: 10.1111/jeb.12814. Epub 2016 Jan 8. J Evol Biol. 2016. PMID: 26688295 Free PMC article.
-
Butterfly Mimicry Polymorphisms Highlight Phylogenetic Limits of Gene Reuse in the Evolution of Diverse Adaptations.Mol Biol Evol. 2019 Dec 1;36(12):2842-2853. doi: 10.1093/molbev/msz194. Mol Biol Evol. 2019. PMID: 31504750 Free PMC article.
-
Pigmentation in Xiphophorus: an emerging system in ecological and evolutionary genetics.Zebrafish. 2014 Feb;11(1):57-70. doi: 10.1089/zeb.2013.0939. Epub 2013 Dec 9. Zebrafish. 2014. PMID: 24320948 Review.
-
The functional basis of wing patterning in Heliconius butterflies: the molecules behind mimicry.Genetics. 2015 May;200(1):1-19. doi: 10.1534/genetics.114.172387. Genetics. 2015. PMID: 25953905 Free PMC article. Review.
Cited by
-
A shared gene but distinct dynamics regulate mimicry polymorphisms in closely related species.bioRxiv [Preprint]. 2025 Mar 3:2025.03.03.641230. doi: 10.1101/2025.03.03.641230. bioRxiv. 2025. PMID: 40093043 Free PMC article. Preprint.
-
Sensitivity of genome-wide tests for mitonuclear genetic incompatibilities.bioRxiv [Preprint]. 2025 Jul 4:2025.06.30.662443. doi: 10.1101/2025.06.30.662443. bioRxiv. 2025. PMID: 40631220 Free PMC article. Preprint.
-
Evolution: A kit for fooling your rivals.Curr Biol. 2024 Oct 21;34(20):R921-R923. doi: 10.1016/j.cub.2024.09.024. Curr Biol. 2024. PMID: 39437728
-
Supergenes are not necessary to explain the maintenance of complex alternative phenotypes.Proc Biol Sci. 2024 Oct;291(2033):20241715. doi: 10.1098/rspb.2024.1715. Epub 2024 Oct 16. Proc Biol Sci. 2024. PMID: 39406344 Free PMC article.
References
-
- Ford EB (1945). Polymorphism. Biol. Rev. 20, 73–88. 10.1111/j.1469-185X.1945.tb00315.x. - DOI
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
