

Semen adaptation to microbes in an insect
- PMID: 39328283
- PMCID: PMC11424074
- DOI: 10.1093/evlett/qrae021
Semen adaptation to microbes in an insect
Erratum in
-
Correction to: Semen adaptation to microbes in an insect.Evol Lett. 2024 Jun 14;8(5):748. doi: 10.1093/evlett/qrae026. eCollection 2024 Oct. Evol Lett. 2024. PMID: 40375869 Free PMC article.
Abstract
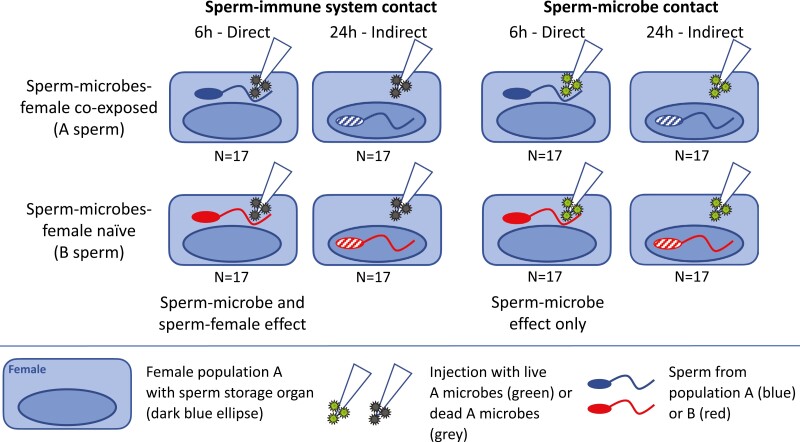
Sperm function is suggested to evolve by sexual selection but is also reduced by microbial damage. Here, we provide experimental evidence that male fertility can adapt to microbes. We found that in vivo, male fertility was reduced by one-fifth if sperm encountered microbes in the females that they had not previously been exposed to, compared to sperm from males that coevolved with these microbes. The female immune system activation reduced male fertility by an additional 13 percentage points. For noncoevolved males, fertility was larger if microbes were injected into females after they had stored away the sperm, indicating microbial protection as a previously unrecognized benefit of female sperm storage. Both medical and evolutionary research on reproductive health and fertility will benefit from considering our findings that the impact of microbes on sperm depends on their joint evolutionary history. Our results may assist in reconciling contradictory results of sexually transmitted disease effects on sperm and bring empirical realism to a recently proposed role of locally adapted reproductive microbiomes to speciation.
Keywords: coevolution; fecundity; host–parasite interactions; speciation.
© The Author(s) 2024. Published by Oxford University Press on behalf of The Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEN).
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Afzelius, B. A., Alberti, G., Dallai, R., Godula, J., & Witalinski, W. (1989). Virus- and rickettsia-infected sperm cells in arthropods. Journal of Invertebrate Pathology, 53(3), 365–377. 10.1016/0022-2011(89)90102-x - DOI
-
- Akaike, H. (1973). Information theory and an extension of the maximum likelihood principle. In Petrov B. N., & Csaki F. (Eds.), Second international symposium on information theory (pp. 267–281). Akademiai Kiado.
-
- Ardia, D. R., Gantz, J. E., Schneider, B. C., & Strebel, S. (2012). Costs of immunity in insects: An induced immune response increases metabolic rate and decreases antimicrobial activity. Functional Ecology, 26, 732–739.
LinkOut - more resources
Full Text Sources
