

Determinants of raffinose family oligosaccharide use in Bacteroides species
- PMID: 39330254
- PMCID: PMC11501099
- DOI: 10.1128/jb.00235-24
Determinants of raffinose family oligosaccharide use in Bacteroides species
Abstract
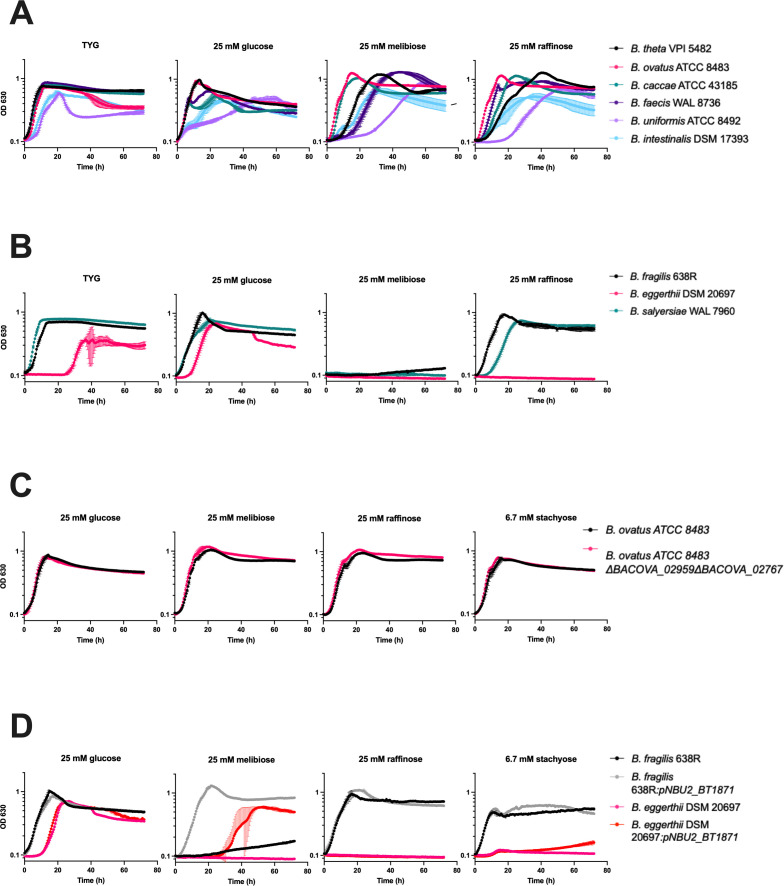
Bacteroides species are successful colonizers of the human colon and can utilize a wide variety of complex polysaccharides and oligosaccharides that are indigestible by the host. To do this, they use enzymes encoded in polysaccharide utilization loci (PULs). While recent work has uncovered the PULs required for the use of some polysaccharides, how Bacteroides utilize smaller oligosaccharides is less well studied. Raffinose family oligosaccharides (RFOs) are abundant in plants, especially legumes, and consist of variable units of galactose linked by α-1,6 bonds to a sucrose (glucose α-1-β-2 fructose) moiety. Previous work showed that an α-galactosidase, BT1871, is required for RFO utilization in Bacteroides thetaiotaomicron. Here, we identify two different types of mutations that increase BT1871 mRNA levels and improve B. thetaiotaomicron growth on RFOs. First, a novel spontaneous duplication of BT1872 and BT1871 places these genes under the control of a ribosomal promoter, driving high BT1871 transcription. Second, nonsense mutations in a gene encoding the PUL24 anti-sigma factor likewise increase BT1871 transcription. We then show that hydrolases from PUL22 work together with BT1871 to break down the sucrose moiety of RFOs and determine that the master regulator of carbohydrate utilization (BT4338) plays a role in RFO utilization in B. thetaiotaomicron. Examining the genomes of other Bacteroides species, we found homologs of BT1871 in a subset and showed that representative strains of species with a BT1871 homolog grew better on melibiose than species that lack a BT1871 homolog. Altogether, our findings shed light on how an important gut commensal utilizes an abundant dietary oligosaccharide.
Importance: The gut microbiome is important in health and disease. The diverse and densely populated environment of the gut makes competition for resources fierce. Hence, it is important to study the strategies employed by microbes for resource usage. Raffinose family oligosaccharides are abundant in plants and are a major source of nutrition for the microbiota in the colon since they remain undigested by the host. Here, we study how the model commensal organism, Bacteroides thetaiotaomicron utilizes raffinose family oligosaccharides. This work highlights how an important member of the microbiota uses an abundant dietary resource.
Keywords: BT1871; Bacteroides; melibiose; polysaccharide utilization loci; raffinose family oligosaccharide; α-galactosidase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Update of
-
Determinants of raffinose family oligosaccharide use in Bacteroides species.bioRxiv [Preprint]. 2024 Jun 7:2024.06.07.597959. doi: 10.1101/2024.06.07.597959. bioRxiv. 2024. Update in: J Bacteriol. 2024 Oct 24;206(10):e0023524. doi: 10.1128/jb.00235-24. PMID: 38895307 Free PMC article. Updated. Preprint.
Similar articles
-
Determinants of raffinose family oligosaccharide use in Bacteroides species.bioRxiv [Preprint]. 2024 Jun 7:2024.06.07.597959. doi: 10.1101/2024.06.07.597959. bioRxiv. 2024. Update in: J Bacteriol. 2024 Oct 24;206(10):e0023524. doi: 10.1128/jb.00235-24. PMID: 38895307 Free PMC article. Updated. Preprint.
-
A Novel Family of RNA-Binding Proteins Regulate Polysaccharide Metabolism in Bacteroides thetaiotaomicron.J Bacteriol. 2021 Oct 12;203(21):e0021721. doi: 10.1128/JB.00217-21. Epub 2021 Jul 12. J Bacteriol. 2021. PMID: 34251866 Free PMC article.
-
Characterization of novel α-galactosidase in glycohydrolase family 97 from Bacteroides thetaiotaomicron and its immobilization for industrial application.Int J Biol Macromol. 2020 Jun 1;152:727-734. doi: 10.1016/j.ijbiomac.2020.02.232. Epub 2020 Feb 21. Int J Biol Macromol. 2020. PMID: 32092418
-
The potential role of nondigestible Raffinose family oligosaccharides as prebiotics.Glycobiology. 2023 May 17;33(4):274-288. doi: 10.1093/glycob/cwad015. Glycobiology. 2023. PMID: 36795047 Review.
-
Significance of Raffinose Family Oligosaccharides (RFOs) metabolism in plants.Adv Biotechnol (Singap). 2024 Mar 19;2(2):13. doi: 10.1007/s44307-024-00022-y. Adv Biotechnol (Singap). 2024. PMID: 39883346 Free PMC article. Review.
References
-
- Karczewski J, Troost FJ, Konings I, Dekker J, Kleerebezem M, Brummer R-J, Wells JM. 2010. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am J Physiol Gastrointest Liver Physiol 298:G851–G859. doi:10.1152/ajpgi.00327.2009 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
